 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
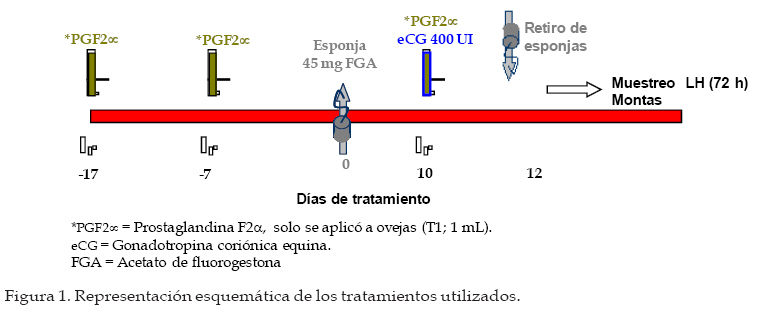
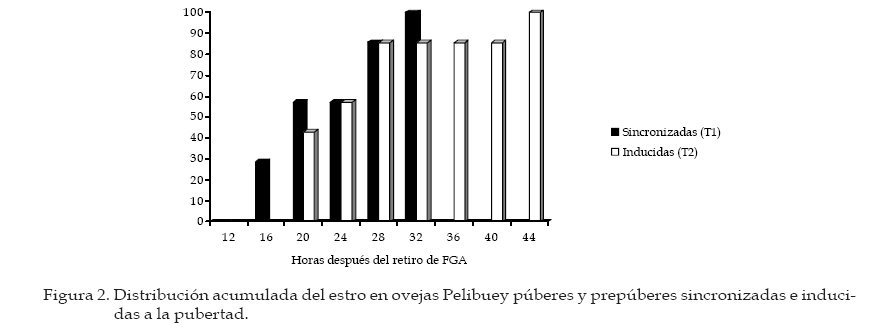
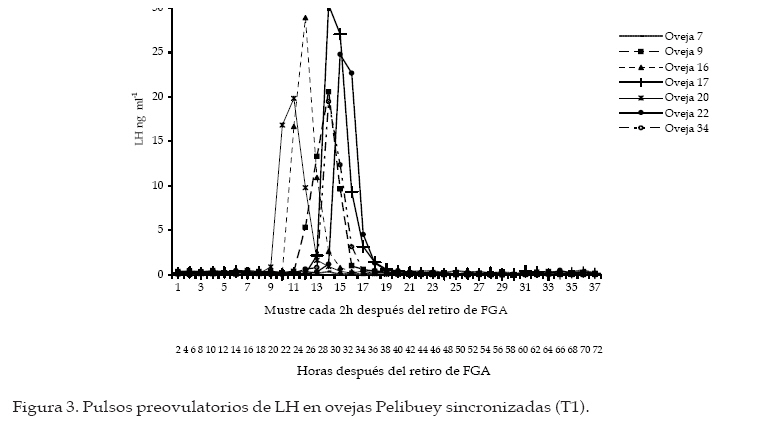
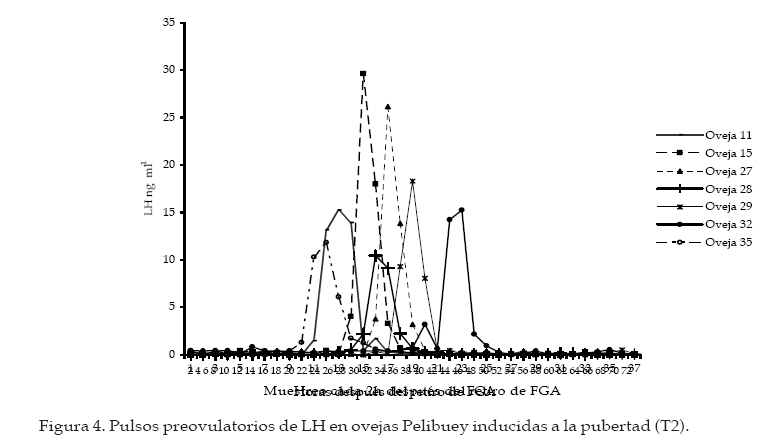
Archivos Latinoamericanos de Producción Animal, Vol. 16, No. 1, 2008, pp. 18-24 Caracteristicas reproductivas de ovejas pelibueysincronizadas e inducidas a la pubertad Reproductive characteristics of pelibuey ewessynchronized and puberty induced J.C. Camacho Ronquillo1*, J. del C. Rodríguez Castillo1, J.E. Hernández Hernández1, A. Pró Martinez2, C.M. Becerril Perez2 y J. Gallegos Sanchez2 1Escuela de Medicina Veterinaria y Zootecnia, Benemérita Universidad Autónoma de Puebla, 4 sur 104 C.P. 72000, Puebla, México. Code Number: la08005 ABSTRACT The aim of the study was to determine the reproductive characteristics of Pelibuey ewes synchronized (T1: n=7), and puberty induced (T2: n=7). Ewes in T1 were pre-synchronized; and seven days after FGA intraviginal sponge were inserted to all ewes during 12 day. Two days prior to sponge removal ewes were injected with eCG. Estrous were observed in 100% of ewes in T1 and T2. Time of estrous was not significant (P>0.05) with values of 21.4 ± 2.2 h and 24.2 ± 3.2 h for T1 and T2. Estrous duration was 60.5 ± 6.6 y 41.3 ± 3.6 for T1 and T2 (P<0.05). Onset, duration and amplitude of LH pre-ovulatory pulse was not significant (P>0.05) with values of 24.5 ± 1.7 and 24.2 ± 4.7 h, 13.4 ± 1.2 and 14.0 ± 0.6 h, 18.1 ± 2.7 and 21.3 ± 3.1 ng mL-1 for T1 y T2 respectively. Pregnancy rate was not significant among treatments with 100% and 85.71% for T1 and T2, respectively. The experiment conclusion was that the used of FGA and eCG in Pelibuey ewes are good alternatives to induce puberty, with acceptable reproductive response. The implementation of these protocols is feasible in AI at fix time programs. Key words: Induction, synchronization, LH pulse, fertility, prolific. RESUMEN El objetivo del experimento fue analizar las características reproductivas de ovejas Pelibuey sincronizadas (T1: n = 7) e inducidas a la pubertad (T2: n = 7). Las ovejas del T1 se presincronizaron, siete días después se aplicó a todas las ovejas FGA intravaginal durante 12 d y dos días antes de retirar el FGA se aplicó eCG. La incidencia de estro fue de 100% en T1 y T2. El inicio del estro no fue diferente (P>0.05) con valores 21.4 ± 2.2 h y 24.2 ± 3.2 h para T1 y T2. La duración del estro fue de 60.5 ± 6.6 y 41.3 ± 3.6 para T1 y T2 (P<0.05). El inicio, duración y amplitud del pulso preovulatorio de LH no mostró diferencias con valores de 24.5 ± 1.7 y 24.2 ± 4.7 h, 13.4 ± 1.2 y 14.0 ± 0.6 h, 18.1 ± 2.7 y 21.3 ± 3.1 ng mL-1 para T1 y T2 respectivamente. La tasa de gestación no fue diferente con 100 y 85.71% para T1 y T2. La tasa ovulatoria y prolificidad fue diferente (P<0.05) con valores de 4.4 ± 1.2 y 2.7 ± 0.4 y 2.5 ± 0.2 y 1.4 ± 0.4, para el T1 y T2 respectivamente. Se concluyó que el uso de FGA y eCG en ovejas Pelibuey es recomendable para inducir la pubertad, ya que las características reproductivas analizadas son aceptables, su implementación puede ser factible en programas de inseminación a tiempo fijo. Palabras clave: Inducción, Sincronización, Pulso de LH, Fertilidad, Prolificidad. Introducción La edad a la pubertad es una variable que afecta rentabilidad del sistema de producción. El periodo directamente la vida productiva de la oveja, tiene re-prepúber esta determinado, principalmente por el lación con la edad al primer parto y por ende con la genotipo y factores como; fotoperíodo, época de nacimiento y la nutrición (I’anson et al., 1997). La inadecuada interacción entre estos factores, provoca diferencias importantes en el inicio de la pubertad, que puede variar desde los 7 meses, hasta cerca de los dos años de edad (Ryan y Foster, 1980). La actividad ovárica esta controlada por el sistema nervioso central, mediante inervaciones noradrenérgicas y peptidérgicas que regulan los receptores a gonadotropinas, estrógenos, progesterona y con la melatonina modifican la frecuencia y amplitud de los pulsos de secreción de GnRH, LH y FSH, esto favorece el crecimiento de folículos preovulatorios que producen estrógenos, hasta llegar a inducir la primera liberación preovulatoria de LH y la primera ovulación (Kinder et al., 1995). Factores como: la edad, el peso, la condición corporal y la época del año, entre otros, pueden ocasionar que no exista sincronía en estos eventos fisiológicos y retrazar el inicio de la pubertad (Goodman, 1994). Sin embargo, avances en fisiología reproductiva ofrecen alternativas para inducir la pubertad, mediante la utilización de hormonas exógenas (Leyva et al., 2000). Actualmente, es necesario caracterizar los eventos fisiológicos que ocurren al inducir y sincronizar el estro, para mejorar la fertilidad. Por lo que, el objetivo de este estudio fue analizar las características reproductivas de ovejas Pelibuey sincronizadas e inducidas a la pubertad, mediante la aplicación de acetato de fluorogestona (FGA) más eCG, bajo el supuesto de que la aplicación de 45 mg de FGA por vía intravaginal durante 12 días más la aplicación de 400 UI de eCG dos días antes de retirar FGA en ovejas prepúberes induce manifestación de estro, ovulación, duración y amplitud del pulso preovulatorio de LH similar al de ovejas púberes sincronizadas. Materiales y Métodos La investigación se realizó en el Colegio de Posgraduados ubicado en Montecillo, Estado de México. Localizado a una latitud norte de 19° 29’, longitud oeste de 98° 53’, a 2250 msnm, el clima es C (W), precipitación media anual de 644.8 mm y la temperatura media anual de 15°C, (García, 1988). Animales Utilizados De un grupo de 34 ovejas Pelibuey, se seleccionaron 14, con buen estado de salud y con una condición corporal de 3.5 a 4 en escala de 1 a 5 Martínez et al. (1999), de estas siete eran púberes (T1) y siete prepúberes (T2) con 236.7 ± 3.1 y 235 ± 0.8 d de edad, 35.5 ± 0.6 y 34.6 ± 1.7 kg de peso respectivamente. Alimentación y Manejo de los Animales Las ovejas se mantuvieron estabuladas, la alimentación fue con avena y alfalfa una vez al día por la mañana y concentrado comercial con 16 % de proteína ofrecido a libre acceso; así como, el agua. Determinación del Grupo Púber y Prepúber Tres semanas antes de iniciar el experimento, se tomaron dos muestras de sangre por semana para analizar la concentración de progesterona. La cual se determinó por radioinmunoanálisis (RIA), en fase sólida con un kit comercial. Se consideró oveja púber si la concentración de progesterona fue mayor a 0.5 ng mL-1 en dos muestras consecutivas ó mayor de 1 ng mL-1 en una sola muestra y prepúberes aquellas que presentaron concentraciones menores de 0.5 ng mL-1 en todas las muestras analizadas (Zarco et al., 1988). Protocolo de Sincronización e Inducción del Estro Con la finalidad de homogenizar la etapa del ciclo estral, las ovejas púberes se presincronizarón mediante la aplicación de dos dosis de PGF2µ, 1 mL por vía intramuscular (7.5 mg de Luprositol) con diez días de intervalo entre aplicaciones; siete días después se aplicó el tratamiento para inducir y sincronizar el estro, el cual consistió en la aplicación de esponjas intravaginales (45 mg de FGA; Chronogest) durante doce días. El día diez del tratamiento, se aplicó 400 UI de eCG (Folligon) por vía intramuscular. Las ovejas del T1 recibieron además 1 mL de PGF2µ. El día 12 del tratamiento se retiraron las esponjas y se inició la fase de toma de muestras de sangre cada dos horas para análisis de LH, la detección de estros y servicios con monta directa (Figura 1). Toma de Muestras de Sangre para Análisis de LH. A partir del retiro de las esponjas se tomaron muestras de sangre cada 2 h, durante 72 h. La determinación de LH se efectúo por radioinmunoanálisis mediante la técnica de doble anticuerpo propuesta por Perera et al. (1992). La sensibilidad de la prueba fue de 0.3 ng mL-1 y el coeficiente de variación fue de 10.26%. Inicio del pulso preovulatorio de LH. Se obtuvo al medir el tiempo transcurrido desde el retiro de FGA, hasta que hubo diferencia significativa (P<0.01) en la concentración de LH y esta se mantuvo en por lo menos dos muestras consecutivas y la concentración de LH fue mayor a 5 ng mL-1. Se consideró que terminó cuando la concentración de LH fue nuevamente basal. Duración del pulso preovulatorio de LH. Se obtuvo la duración individual mediante un análisis de mediciones repetidas en un diseño completamente al azar Littell et al. (1998), posteriormente, con los resultados individuales se realizó una comparación de medias. Amplitud del pico preovulatorio de LH. Se obtuvo de manera individual, al restar a la máxima concentración de LH, la concentración basal, esta ultima se obtuvo al promediar la concentración de LH en los periodos que se encuentran antes y después del pulso preovulatorio de LH Mattioli et al. (1986), posteriormente, se efectúo una prueba de comparación de medias. Detección del Estros y monta natural. Inició a partir de 12 h de retirado el FGA, se efectuo cada 2 h con ayuda de cuatro machos provistos de un mandil. Las ovejas en estro recibieron dos montas, al inicio del estro y 12 h después. Duración del Estro: Se considero el tiempo desde el inicio del estro hasta que la oveja dejó de ser receptiva al macho, la detección fue cada dos horas por lapsos de 20 min. Tasa de Ovulación: Se efectuó por conteo de cuerpos lúteos, 10 d después de la monta mediante laparoscopia. Diagnóstico de Gestación: Se realizó mediante ultrasonografía, 45 d después de la monta. Análisis Estadístico: Para determinar la duración y amplitud del pulso preovulatorio por ovejas, se efectuó un análisis estadístico con el procedimiento MIXED de SAS Littell et al. (1999), para un modelo estadístico de mediciones repetidas en un diseño experimental completamente al azar, considerando el periodo como variable fija y la concentración de LH como aleatoria. Los datos obtenidos para las variables; inicio del estro, duración del estro, inicio del pulso preovulatorio de LH, duración del pulso preovulatorio de LH, amplitud del pulso preovulatorio de LH y prolificidad se analizaron mediante una prueba de "t" de Student , para un diseño completamente al azar. La prueba se realizó con el procedimiento denominado TTEST del programa (SAS Institute, 1999). Para analizar las variables incidencia de estros y tasa de gestación se realizó una prueba de Ji-cuadrada. Resultados y discusión Incidencia de Estros No se observó diferencia (P>0.05), el 100% de los animales de T1 y T2 manifestaron estro. Esto evidencia la efectividad de los progestágenos y eCG que puede ser de 95 a 100% (Molina et al., 2005). Otros investigadores señalaron que la aplicación de FGA por periodos de 12 a 14 d, ocasionaron resultados similares en época reproductiva y en anestro estacional Robinson (1988), esto explica la respuesta en la inducción de la pubertad. Rosado et al. (1998), mediante el esquema de sincronización denominado INRA, similar al de esta investigación obtuvieronel 97% de incidencia de estros, Ávila et al. (1997) reportaron 90% de estros con el uso de FGA más la aplicación de 500 UI de eCG. Inicio del Estro No se encontró diferencia (Figura 2; P>0.05). El inicio del estro, se presentó a las 21.4 ± 2.2 h y a las 24.2 ± 3.2 h, en T1 y T2. Esto posiblemente debido al efecto de eCG en la liberación de FSH y LH, que favorece la retroalimentación positiva con estradiol, como lo mencionó Rosado et al. (1998). En esta investigación la presentación temprana del estro, se atribuyó a la aplicación de eCG dos días antes de retirar el FGA; ya que, favorece la producción de estrógenos por el folículo preovulatorio y concuerda con resultados obtenidos por (Rangel et al., 1993 y Ungerfeld y Rubianes, 1999). Sin embargo, Molina et al. (2005), indicaron que el estro se presenta en un intervalo de 32 a 36 h con administración de eCG al momento de retirar las esponjas. Otro factor que disminuye el tiempo desde el retiro de FGA hasta el inicio del estro; así como, su dispersión, es la presencia intermintente del macho utilizado en la detección de estros, como lo demostraron Ungerfeld y Rubianes (1999) y en el presente estudio pudo contribuir para que la distribución del estro fuera similar en ambos tratamientos. Duración del Estro Se observó diferencia entre tratamientos (P<0.05) con valores de 60.5 ± 6.6 y 41.3 ± 3.6 h para T1 y T2. Gordon, (1997) y Fernández et al. (1997), señalaron que la duración del estro en ovejas varía de 24 a 48 h. Sin embargo, puede aumentar hasta en un 50% en ovejas con ovulación múltiple. Además, González et al. (2000) indicaron que la duración del estro puede disminuir a la mitad cuando las hembras permanecen con el macho con respecto a ovejas en las que se detecta el estro con el macho, de manera intermitente cada 2 h, como se efectúo en este experimento. Se ha demostrado la participación de estradiol y GnRH en el control de la conducta estral. El pulso preovulatorio de LH es consecuencia de la liberación preovulatoria de GnRH, este se origina por un mecanismo de retroalimentación positivo con el estradiol que es liberado por el folículo preovulatorio y actúa en el hipotálamo medio basal para desencadenar el proceso que culmina con la ovulación, la concentración de estradiol esta relacionado con la magnitud de la liberación de GnRH (Caraty et al., 1999). El mayor número de folículos en T1 posiblemente sea la causa de mayor duración del estro. Inicio del Pulso Preovulatorio de LH No se observó diferencia entre tratamientos (Figura 3 y 4; P>0.05) el inicio fue a las 24.5 ± 1.7 y 24.2 ± 4.7 h, para T1 y T2. Estos resultados difieren de lo reportado por Robinson (1988) y López-Sebastián (1991) quienes señalaron que el pico preovulatorio de LH inicia de 36 a 39 h después de retirar el progestágeno. En este estudio, este resultado pudo ocasionarse debido a que se aplicó eCG dos días antes de retirar FGA. Phogat et al. (1999), indicaron que la concentración elevada de estradiol reduce el tiempo en la presentación del pulso preovulatorio de LH. La aplicación de 50 µg de estradiol al retirar el progestágeno, resulta en un inicio del pulso a 20.5 ± 2.0 h. El efecto del estradiol sobre la inducción del pulso preovulatorio de LH y comportamiento sexual se observa en la oveja ovariectomizada durante la estación reproductiva y en anestro (Goodman et al., 1994). De este modo el pico preovulatorio de estradiol llega al área preóptica del hipotálamo donde regula la formación de los pulsos preovulatorios de LH y FSH y por ende la ovulación (Caraty et al., 1999). Duración del Pulso Preovulatorio de LH No se observó diferencia (Figuras 3 y 4; P>0.05), los valores fueron 13.4 ± 1.2 y 14.0 ± 0.6 h para T1 y T2. Molina et al. (2005) reportaron resultados similares con duración de 13.3 y 14 h (P>0.05) en ovejas con y sin cuerpo lúteo previo al tratamiento con progestágeno. Dobson et al. (1999), señalaron que la duración del pico fue 8.4 ± 0.9 h con uso de estradiol. Estas investigaciones muestran que el fármaco utilizado en la sincronización influye en la duración del pulso preovulatorio de LH. Amplitud del pico preovulatorio de LH. No hubo diferencias entre tratamientos (P>0.05), la concentración media de LH fue de 18.1 ± 2.7 y 21.3 ± 3.1 ng mL-1 para T1 y T2 respectivamente, y son similares a lo reportado por Dobson et al. (1999), quienes señalaron una amplitud de 18.4 ± 0.9 ng mL-1. Harris et al. (1999) indicaron una amplitud de 5.9 ± 0.7 ng mL-1 después del uso de estradiol. Tasa Ovulatoria El número de cuerpos lúteos fue diferente (P<0.05) con valores de 4.4 ± 1.2 y 2.7 ± 0.4 para el T1 y T2. Se sabe que, desde el periodo prepúber se encuentran folículos ováricos sensibles a LH que requieren una exposición previa a progesterona para que la calidad y el número de éstos aumente (Caraty et al., 1993; Campbell et al., 1999, Leyva et al., 2000). No obstante, aunque las ovejas prepúberes pueden responder con ovulaciones después de administrar gonadotropinas exógenas, la tasa ovulatoria es inferior que el de ovejas púberes (Dobson et al., 1999; Gonzales et al., 2000). Las investigaciones indican que se obtienen mejores resultados al aplicar eCG dos días antes de retirar el progestágeno con lo que puede lograrse hasta 4.5 CL (Rangel et al., 1993), similar a lo encontrado en el T1. El número de ovulaciones en ambos grupos de ovejas se atribuye al efecto de eCG que estimula el crecimiento folicular y disminuye la atresia folicular con lo que aumenta el número de folículos ovulatorios (Caraty y Skinner, 1999). Tasa de Gestación El diagnóstico de gestación mostró que el 100% de las ovejas del T1 estuvo gestante, mientras que el T2 obtuvo el 85.71% (P>0.05). En esta investigación el pequeño número de repeticiones pudo ocasionar que no se encontraran diferencias estadísticas. Investigaciones previas sugieren que el mayor porcentaje de fertilidad se obtiene en las ovejas púberes debido a la mayor cantidad de folículos capaces de ovular, liberar un oocito normal y formar un cuerpo lúteo de vida media normal (Crosby et al., 1991; Campbell et al., 1999). Prolificidad El tamaño de la camada obtenido mostró diferencia (P<0.05), los resultados fueron 2.5 ± 0.2 y 1.4 ± 0.4 crías, para el T1 y T2. Para el T1 es superior a lo reportado por Rosado et al. (1998) González et al. (2000), quienes reportaron 2.1 corderos por oveja, después de aplicar progestágeno y 500 UI de eCG. El resultado del T2 aunque con media menor, es aceptable; ya que en la inducción de la pubertad, la tasa de ovulación y prolificidad son generalmente inferiores al de ovejas con actividad reproductiva (Godman, 1994). Sin embargo, el resultado obtenido es mayor a la presentan las ovejas Pelibuey. González et al. (2000), reportan de 1.1 a 1.4 de prolificidad. Conclusiones De acuerdo con los resultados obtenidos se reco-de inseminación a tiempo fijo debido baja dispersión mienda sincronizar el estro e inducir la pubertad en de presentación de estro y al alto grado de sincronía ovejas Pelibuey con FGA por 12 d y eCG dos días de la presentación del pulso preovulatorio de LH, lo antes de retirar FGA, ya que las ovejas prepúberes que puede favorecer el encuentro entre el óvulo y el obtienen variables reproductivas aceptables. El tra-espermatozoide en el sitio de fecundación, y optimizar tamiento hormonal puede utilizarse en programas la fertilidad. Literatura citada
© 2008 ALPA. Archivos Latinoamericanos de Producción Animal The following images related to this document are available:Photo images[la08005f4.jpg] [la08005f2.jpg] [la08005f1.jpg] [la08005f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}