 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
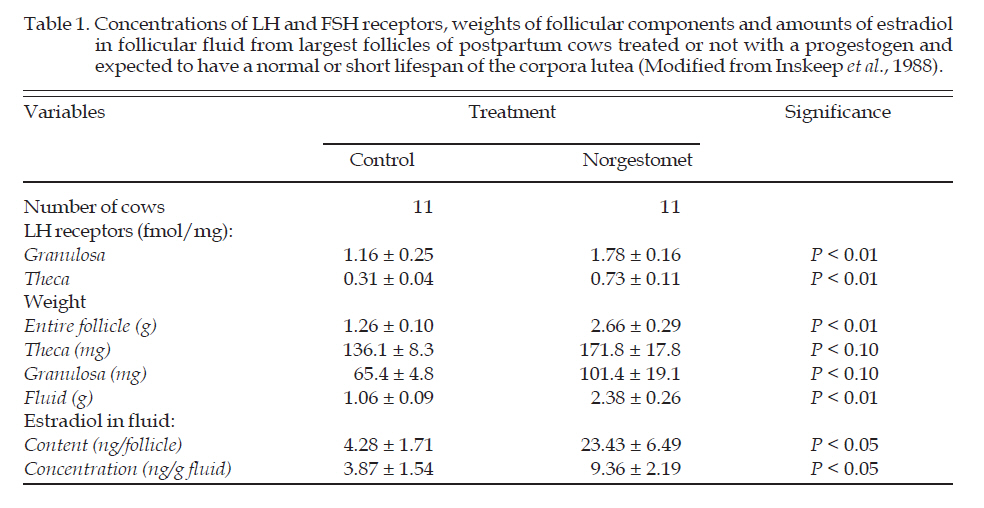
Archivos Latinoamericanos de Produccion Animal, Vol. 16, No. 3, July-September, 2008, pp. 175-185 Original Article Infertilidad asociada con la duración de la fase luteal en vacas postparto F Perea G1, E Keith Inskeep2 1 Departamento de Ciencias Agrarias, Universidad de Los Andes, Trujillo, Venezuela Correspondence Address: F Perea G Departamento de Ciencias Agrarias, Universidad de Los Andes, Trujillo Venezuela ferromi@ula.ve Date of Submission: 12-Dec-2007 Code Number: la08023 Abstract After first postpartum ovulation most cows have a short luteal phase due to a premature secretion of PGF2α from the uterus. Pregnancy rate in estrous cycles with short luteal phases is essentially nil. The lack of exposure of the uterus to P4 and estrogen prior to first postpartum ovulation decreases the concentration of P4 receptors and up regulates oxytocin receptors. The inadequate number of P4 receptors and presence of endometrial oxytocin receptors earlier in cows with short than normal luteal phases allows premature release of PGF2α and the resultant regression of the CL. During luteolysis, uterine and luteal PGF2α cause death of the embryos mainly during the transition from morula to blastocyst when the embryos are more susceptible to embryotoxic effect.Keywords: postpartum, first ovulation, short luteal phase, infertility, cattle Resumen Luego de la primera ovulación postparto, la mayoría de las vacas presentan un ciclo estrual corto debido a la secreción anticipada de PGF2a desde el útero. La tasa de preñez en ciclos estruales con una fase luteal corta es prácticamente cero. La falta de exposición del útero a la P4 y estrógenos antes de la primera ovulación postparto, disminuye la concentración de receptores de P4 y estimula la expresión de receptores de oxitocina. El número inadecuado de receptores uterinos de P4 y la presencia de receptores endometriales de oxitocina mas temprano en vacas con ciclos estruales cortos que con ciclos estruales normales, permite la secreción prematura de PGF2a y la resultante regresión del cuerpo lúteo. Durante la luteólisis, la PGF2a de origen uterino y luteal causan la muerte embrionaria principalmente durante la transición de morula a blastocito, cuando los embriones son mas susceptibles al efecto embriotóxico. Palabras clave:postparto, primera ovulación, fase luteal corta, infertilidad, ganado vacuno. Introduction The principal goal of cattle operations is to achieve calving and lactation once every year, which allows maximum expression of biological productivity, leading to greatest milk and beef sales (Perea et al., 2002). In order to accomplish this goal, conception should occur before day-90 postpartum. However, several environmental, genetic, and physiologic factors can contribute to low fertility and embryonic losses once cows reinitiate reproductive cyclicity (Perea et al., 2005). The postpartum interval is a transition period during which the reproductive axis recovers after gestation to progressively reach the anatomical and functional status needed to initiate a new pregnancy. The presence of dominant follicles in the ovaries, as well as an adequate pattern of LH secretion, leads to the first postpartum ovulation a few weeks after parturition (Murphy et al., 1990; Rajamahendran and Taylor, 1990; Perea et al., 1998). Fertility at this first estrual cycle is lower than in subsequent cycles, mainly attributable to embryonic death during the first two weeks after first insemination (Wells et al., 1985; Inskeep, 2002). Premature secretion of PGF2α and early regression of the corpus luteum (CL) are responsible for the reduced conception rate (Cooper et al., 1991; Zollers et al., 1993; Inskeep and Dailey, 2004). Thus, maternal recognition of pregnancy is prevented, and a few days later the onset of a subsequent estrous cycle, which in turn has a normal conception rate (Odde et al., 1980; Ramirez-Godinez et al., 1981). In fact, as will be documented in this review, the premature secretion of PGF2α in the first cycle is embryotoxic and the embryo dies even if exogenous progesterone is provided. Short estrous cycles with sub-functional corpora lutea (CL) are frequent before the first ovulatory estrus in peripubertal heifers (Berardinelli et al., 1979), in postpartum dairy (Menge et al., 1962; Morrow et al., 1966) and beef (Corah et al., 1974) cows and as a consequence of early weaning of beef calves (Bellows et al., 1974; Odde et al., 1980). Treatment with progesterone (P4) prior to the first ovulation induces a CL with normal lifespan (Ramírez-Godinez et al., 1981; Zollers et al., 1993; Fike et al., 1997) in a large proportion of cows. An experimental model for comparisons of normal or short estrous cycles at the same stage postpartum was developed based on these facts. For short cycles, either early weaning or treatment with GnRH, hCG or estradiol will suffice; whereas for normal estrous cycles, these same treatments can be preceded by treatment with a progestogen. The model has been used extensively to study the mechanisms involved in short cycles following the first postpartum ovulation (Shefffel et al., 1982; Buford et al., 1996, Hernandez-Fonseca et al., 2000), as well as to find therapeutic alternatives to improve reproductive efficiency during postpartum anestrus (Butcher et al., 1992; Breuel et al., 1993; Schrick et al., 1993; Buford et al., 1996; Fike et al., 1997). Characteristics of postpartum cattle Resumption of ovarian cyclicity in Bos taurus dairy and beef cows takes place by around 30 or 60 days following parturition, respectively (Roche et al., 1992), although this period is longer in tropical crossbred cows (Bos taurus x Bos indicus) (Perea et al., 2002, 2005; Gonzalez et al., 2003). The occurrence of a short interovulatory interval (7 to 12 days) is frequent after first postpartum ovulation (Macmillan et al., 1971; Odde et al., 1980; Ramirez-Godinez et al., 1982; Copelin et al., 1989; Zollers et al., 1989; Cooper et al., 1991; Zollers et al., 1993). The incidence of short estrual cycles reported in postpartum cows ranged from 7 to 100% (Macmillan et al., 1971; Odde et al., 1980; Corah et al., 1974; Wells et al., 1985; Murphy et al., 1990; Savio et al., 1990; Stagg et al., 1995; Mackey et al., 2000). However, in cows induced to ovulate by early weaning or treatment with GnRH or hCG, the occurrence ranged between 60 and 80% (Odde et al., 1980; Kesler et al., 1980; Ramírez-Godinez et al., 1981; Copelin et al., 1988; Johnson et al., 1992). Fertility associated with first estrous cycles was lower than in subsequent cycles, which had normal lifespan of the CL (Odde et al., 1980; Ramirez-Godinez et al., 1982; Breuel et al., 1993). Physiologic bases of short luteal phase In initial studies, short luteal phase was not associated with either decreased luteotropic support by LH (Garcia-Winder et al., 1986; Copelin et al., 1988) or greater luteal sensitivity to PGF2α (Copelin et al., 1988; [Figure - 1]). Non-suckled hysterectomized cows weaned at 23 to 30 days postpartum did not return to estrus following first estrus, while intact cows had estrual cycles averaged 8.8 days (Copelin et al., 1987; [Figure - 2]). Although concentrations of P4 were similar from estrus to day 5, secretion of P4 decreased after day 5 of the first cycle, but not after second estrus in intact cows or following the first or a second estrus (induced by PGF2α) in hysterectomized cows (Copelin et al., 1987; [Figure - 1] and [Figure - 2]). Patterns of P4 before and after day 5 postestrus in postpartum cows expected to have CL of normal or short lifespan had similar profiles (Zollers et al., 1989, 1991, 1993). Active immunization of anestrous beef cows against PGF2α extended the lifespan and secretory capacity of CL (Copelin et al., 1989). The physiologic phenomenon has been described as a subnormal luteal phase occurring due to a premature release of PGF2α from the uterus and concomitant early CL regression (Copelin et al., 1987; Cooper et al., 1991). Presumably, lack of P4 caused death of the embryo and the onset of a new estrual cycle. During normal estrous cycles, luteal phase P4 allows a rise in estrogen to promote synthesis of P4 receptors (Zollers et al., 1993) and to inhibit synthesis of uterine receptors for oxytocin through the midluteal phase (McCracken et al., 1984; Kieborz-Loos et al., 2003). However, the lack of exposure of the uterus to P4 followed by estrogen prior to first postpartum ovulation decreases the concentration of P4 receptors and up regulates oxytocin receptors, which allows premature secretion of PGF2α (Cooper et al., 1991). Although embryonic death was presumably due to a lack of P4 support, exogenous P4 did not prevent embryonic death, and evidence began to accumulate that PGF2α might act as an embryotoxin (Butcher et al., 1992; Breuel et al., 1993). During luteolysis both uterine and luteal PGF2α cause death of the embryos mainly during the transition from morula to blastocyst when the embryos are more susceptible to embryotoxins (Buford et al., 1996; Seals et al., 1998; Hockett et al., 2004). Furthermore, the process of embryo transfer stimulates uterine secretion of PGF2a and success rates have been improved by treatment with inhibitors of prostaglandin synthesis at or near the time of transfer (Scenna et al., 2005). Mean concentrations of PGF2α in vena caval plasma on days 4 to 9 after injection of hCG were greater in postpartum beef cows undergoing premature luteal regression than in those with a normal luteal phase (Cooper et al., 1991). Peter et al. (1989) found increased concentrations of oxytocin in association with increased concentrations of PGFM in dairy cows with short luteal phases, but Cooper et al. (1991) did not find consistent increases in oxytocin in association with increases in PGF2α in plasma from the inferior vena cava of beef cows. Daily administration of oxytocin from day 2 postestrus in beef cows markedly reduced length of the estrous cycle, but it did not appear to influence the rate at which P4 concentration increased during the first days after estrus (Donaldson et al., 1970). In postpartum cows, oxytocin induced a release of PGF2α on day 5 of a short but not a normal estrous cycle, and the oxytocin-induced release of PGF2α on day 5 of the short estrous cycle was similar to that near to the time of luteolysis (day 16; Zollers et al., 1989). Moreover, in vitro secretion of PGF from endometrial explants on day 5 was greater for cows anticipated to have a short than a normal luteal phase (Zollers et al., 1991). Concomitantly, endometrial concentration of oxytocin receptors was greater and mean number of P4 receptors per cell was lower on day 5 after estrus in postpartum beef cows expected to have short luteal phase [Figure - 3] than in cows receiving P4 pre-treatment and expected to have a normal luteal phase (Zollers et al., 1993). Although P4 pre-treatment is necessary for normal luteal lifespan and timing of uterine secretion of PGF2α following first ovulation in cattle, early evidence indicated that oxytocin may be involved in premature secretion of PGF2α in cows with short estrual cycles. Luteolysis during the estrous cycle in ruminants was believed to occur as consequence of a reciprocal positive feedback mechanism between luteal oxytocin and uterine PGF2α (McCracken et al., 1984; Lafrance and Goff, 1988; Lamsa et al., 1989). This mechanism was proposed to be responsible for the episodic secretion of PGF2α necessary to complete luteal regression (Flint et al., 1990); furthermore, this process only occurs at a specific time of the estrous cycle (Lafrance and Goff, 1988; Silvia et al., 1991; Kieborz- Loos et al., 2003). It was believed that P4 suppressed secretion of PGF2α when more P4 receptors were present, presumably by inhibition of synthesis of oxytocin receptors early in the estrous cycle when the uterus was more responsive to P4 (McCracken et al., 1984; Hazzard et al., 1998; Kieborz-Loss et al., 2003). However, as the estrous cycle progressed, concentrations of P4 receptors decreased, which in turn allowed the synthesis of estrogen and oxytocin receptors (McCracken et al., 1984). As a result, the uterus became more responsive to oxytocin and a positive feedback mechanism was apparently established between oxytocin and endometrial PGF2α. In the normal estrous cycle of ruminants, a period of progestational influence appears to be important in the timing of luteal regression. Treatment with exogenous P4 on day 0 or 1 after estrus advanced the occurrence of the first peak of PGF2α and shortened the length of the estrous cycle in sheep (days 8 and 11 for P4 -treated ewes and days 12.3 and 17 for control ewes, respectively; Ottobre et al., 1980). In addition, the blockade of P4 receptors by mifepristone (RU 486) during the early to mid-luteal phase inhibited luteolysis and preserved a functional corpus luteum beyond day 20, presumably due to insufficient PGF2α secretion from the endometrium (Morgan et al., 1993). Abolition of P4 source on days 12 or 14 by removal of either ovaries or CL in sheep, resulted in a temporal increase of PGF2α followed by a gradual decline (Ottobre et al., 1984; Vincent et al., 1986), suggesting that continued secretion of P4 is necessary for secretion of PGF2α to continue beyond day 14 of the estrous cycle. Apparently, P4 regulates secretion of PGF2α by timing the initial peaks of secretion early in the estrous cycle, and thereafter, by modulating secretion of PGF2a until luteal regression is completed. However, in postpartum cows and anestrous ewes the absence of a previous exposure of the uterus to P4 followed by estrogen prior to first ovulation alters the physiological conditions of the uterus and as a consequence, the timing of luteolysis. Endocrine events conducive to short or normal luteal phase Preovulatory follicles from anestrous beef cows pre-treated with the synthetic progestogen, norgestomet, were larger and contained more estradiol-17b than those of control cows with short estrous cycles (Sheffel et al., 1982; Garcia-Winder et al., 1986). Furthermore, greater concentrations of receptors for LH, in both granulosa and theca, and greater concentrations of estradiol were detected in the largest preovulatory follicles collected from postpartum cows treated with norgestomet than from control cows (Inskeep et al., 1988; [Table 1]). In addition, postpartum anestrous cows treated with a subcutaneous ear implant containing estradiol-17b for 40 days after calving had approximately 40% fewer short estrous cycles during the time of estradiol exposure than non-treated cows (Day et al., 1990). Increased concentrations of estradiol have been associated with induction of endometrial P4 receptors (Zelinski et al., 1980; Ing et al., 1997). Therefore, lower concentrations of estradiol secreted by preovulatory follicles from cows that would have a short luteal phase (Sheffel et al., 1982; Garcia-Winder et al., 1986) could reduce uterine P4 receptor synthesis (Zollers et al., 1993) and consequently the first rise in P4 is followed by early secretion of PGF2α (Cooper et al., 1991), which shortened the luteal phase. A confirmatory study of previously documented findings was carried out by Kieborz-Loos et al. (2003). Postpartum anestrous cows exposed or not to different combinations and/or sequences of exogenous estradiol-17b and P4 were ovariectomized on day 23 or 32 after calving. Oxytocin was injected at the end of each treatment to test the ability of the postpartum uterus to secrete PGF2α. Oxytocin induced secretion of PGF2α was greater on day 16 than on day 6 of a simulated normal luteal phase, implying that steroid treatment (P4 followed by estradiol) stimulated the uterine responsiveness to oxytocin similar to that observed at end of a normal estrous cycle (Lafrance and Goff, 1988). However, neither P4 alone nor estradiol-17b followed by P4 treatment were capable of reducing PGF2α secretion on day 6 of a simulated estrual cycle in ovariectomized cows. Only when estradiol-17b/P4 treatment was preceded by a period of P4 priming, exogenous oxytocin was unable to provoke an increased secretion of PGF2α on day 6 of the simulated cycle (Kieborz-Loos et al., 2003). In postpartum anestrous cows, comparatively more cows ovulated and formed CL with typical lifespan after treatment with a P4 releasing intravaginal device alone or in combination with estradiol benzoate (EB) than when cows were treated with only EB or were untreated controls (C) (Fike et al., 1997). Interestingly, the addition of EB enhanced the proportion of cows with normal length of the estrual cycle (P4 +EB: 76%; P4 : 57%; EB: 24%; C: 19%). What is the source of the steroids that assure a normal lifespan of the CL in those cows during spontaneous ovulatory cycles after calving? In prepubertal heifers, Berardinelli et al. (1979) showed that small luteal tissue embedded in the ovarian stroma and surrounded by connective tissue were responsible for increases in P4 before first normal ovulatory cycles. In addition, a gradual rise in P4 prior to the first postpartum estrus has been reported in cattle (Donaldson et al., 1970; Corah et al., 1974; Perry et al., 1991; Dimmick et al., 1991; Breuel et al., 1993). Increased P4 3 days before first postpartum estrus was detected in primiparous cows that conceived, while no elevation prior to estrus was detected in those cows that failed to conceive and had a short estrous cycle (Corah et al., 1974). Also, Ramirez-Godinez et al. (1981) reported that 7 cows with estrous cycle of at least 17 days had a significant rise in serum progesterone prior to the first detected estrus. The occurrence of ovulations in absence of clinical signs of estrus frequently observed in postpartum cows spontaneously reinitiating ovarian activity might account for the particular sequence of endocrine events required for a normal luteal phase (Stevenson and Britt, 1979; Fike et al., 1997; Henao et al., 2000). In this regard, Castenson et al. (1976) reported that the increase in P4 prior to first postpartum estrus was produced by a short lived CL, presumably from ovulation because of the presence of a stigma. A more recently study using ultrasonography supported those findings (Perry et al., 1991). Fertility and embryo viability in cows with short luteal phase As reviewed above, the first spontaneous or induced ovulation after calving is associated with a CL of short lifespan (for review see: Inskeep, 1995; 2002; Inskeep and Dailey, 2004). Even when first ovulation is preceded by estrus, fertility in beef cows is lower than in first normal length or subsequent estrous cycle (Ramirez-Godinez et al., 1981; Breuel et al., 1993). Fertilized oocytes in different stages of development were recovered from oviduct and uterus (on days 3 and 6 after first estrus, respectively) from cows with a short-lived CL (Ramirez-Godinez et al., 1982; Breuel et al., 1993; Schrick et al., 1993). The failure to sustain pregnancy was presumed to be due to early luteal regression, as indicated by serum concentrations of P4 (Ramirez-Godinez et al., 1982). Surprisingly, neither daily provision of melengestrol acetate nor twice daily injection of P4 from day 4 to 35 after estrus, maintained pregnancy in cows with short-lived CL (Breuel et al., 1993). Thus, premature regression of the CL was not the only factor responsible for the low fertility associated with the short luteal phase. Two good-quality embryos were transferred on day 7 into cows induced to have short or normal luteal phases and supplemented with daily doses of P4 from day 4 to 35. Pregnancy rates were 28% and 56% for short and normal luteal phases, respectively (Butcher et al., 1991). In a later study, uterine horns ipsilateral to the CL from cows induced to have short or normal luteal phases were flushed nonsurgically and embryos were transferred into recipient cows with normal estrous cycles (Schrick et al., 1993). Quality and stage of development of embryos on day 6 did not differ among groups. Pregnancy rate did not differ after embryos were transferred into cycling recipients (50 vs 72% for embryos from short and normal cycles, respectively). However, on the basis of donor animals, only 23% of recovered embryos from short cycles were ultimately successful pregnancies compared to 46% from normal cycles. Apparently, both detrimental factors that affected the oocytes or embryos before day 7 and an improper uterine environment after day 7 could contribute to early embryonic death (Butcher et al., 1991; Schrick et al., 1993). Embryotoxic effect of prostaglandin F2α Inskeep (1995) stated that subluteolytic concentrations of PGF2α could play a role in embryonic death during early development via release of an embryotoxin from the CL. In characterizing such embryotoxic factors, Schrick et al. (1993) found concentrations of PGF2α in uterine flushings to be greater in cows with short than normal luteal phases on day 6; the most relevant finding was that the more PGF2α in the uterine fluid the lower the quality of the embryos (r = -0.42; P<0.07). Therefore, a detrimental uterine environment appeared to be a major cause of low fertility in cows with short luteal phases after first ovulation (Schrick et al., 1993). A series of experiments was designed to study the role of the PGF2α in reducing fertility in cows with short luteal phases. Cycling cows were injected every 8 hours with saline (control), PGF2α or flunixin meglumine (an inhibitor of prostaglandin endoperoxide synthetase), on days 4 through 7 or 5 through 8, and supplemented with exogenous P4 twice daily from day 3 until pregnancy diagnosis was performed. In one group of PGF2α-treated cows, the CL was removed on day 7. As expected, pregnancy rate was lower in cows treated with PGF2α compared with those that received saline or flunixin meglumine [Figure - 4], but surprisingly, pregnancy was restored to normal when treatment with PGF2α was accompanied by lutectomy (Buford et al., 1996). Cows induced to have short estrous cycles by weaning at 25 to 31 days postpartum were treated with saline (S), flunixin meglumine (F), flunixin meglumine plus lutectomy or lutectomy only (L), on days 4 through 9, and supplemented with exogenous progesterone twice daily from day 3 until pregnancy diagnosis. Pregnancy rate did not differ among S, F and L groups (< 21%). However, pregnancy rate was higher (50%; [Figure - 4]) when lutectomized cows were treated with flunixin meglumine (Buford et al., 1996). That treatment would have reduced secretion of PGF2α by the uterus and removed the CL as a source of PGF2α. Moreover, in ewes treated with PGF2α on days 4 through 7 after mating and supplemented with an intravaginal progestogen, lutectomy increased pregnancy rate (59%) compared with sham lutectomy (33%; Costine et al., 2001). In summary, lutectomy and the inhibition of prostaglandin synthetase prevented the negative effects of endogenous PGF2α on the early embryonic stage in postpartum cows (Buford et al., 1996), whereas lutectomy alone was effective in nonlactating cycling cows (Buford et al., 1996), and ewes (Costine et al., 2001) treated with exogenous PGF2α. Therefore, the lower embryo survival in postpartum cows appeared to be due to the combined effect of uterine PGF2α secreted on days 4 through 9 (Cooper et al., 1991) and the luteal secretion of PGF2α in response to the uterine signal (Buford et al., 1996). In subsequent studies pregnancy rate was lowered by daily dosage with PGF2α, on day 5 through 8 (23%) but not on days 10 to 13 (50%) or 15 to 18 (60%) after mating of P4 -supplemented cows (Seals et al., 1998). Treatment of P4 -supplemented cows with PGF2α on day 5 through 8 reduced quality and retarded development of embryos (Hockett et al., 2004). Also, PGF2α in vitro reduced the ability of 26 to 32-cell bovine embryos to develop to blastocysts (Scenna et al., 2004). Therefore, PGF2α may interfere with the process of compaction of the embryo at the morula stage (Hockett et al., 2004), probably by alterations in gap junctions and cell adhesion molecules, induction of apoptosis mechanisms or alterations of gene transcription during embryonic development (Scenna et al., 2004). In order to test whether the luteal source of the embryotoxin was delivered by a systemic or a local pathway, beef heifers and cows received one good quality embryo into each uterine horn on day 6 or 7 of the estrual cycle, and P4 replacement therapy from transfer through day 35 (Hernandez-Fonseca et al., 2000). Each female was injected with three daily therapeutic doses of PGF2α or saline from day 5 through 8. Survival of embryos placed into the uterine horn adjacent or opposite to the CL was not different and no evidence was found to support the existence of a local embryotoxic effect originated from the regressing CL. Therefore, embryotoxicity probably involved compounds that either act systemically or were transported via the uterine lumen to the uterine horn contralateral to regressing CL (Hernandez- Fonseca et al., 2000). In a similar experiment in sheep, the embryotoxic effect of the regressing CL also was demonstrated; again, there was evidence that the effect occurred through a systemic pathway (Costine et al.,2001). Is oxytocin involved as an embryotoxic component of the regressing corpus luteum? At the time of Inskeep′s review in 1995, oxytocin was included in the list of factors that could play a role in embryonic death in cows with short luteal phases. As was previously pointed out, luteal oxytocin and uterine prostaglandin were believed to participate in luteolysis through a reciprocal positive feedback mechanism (McCracken et al., 1984). Concentrations of endometrial oxytocin receptors were greater at day 5 after estrus in cows with short lifespan of the CL than in cows receiving P4 pretreatment and expected to have a normal luteal phase (Zollers et al., 1993). Greater concentrations of oxytocin were evident 30 minutes after first injection of PGF2α in P4 -supplemented cows treated three times daily with PGF2α on day 5 to 8 after estrus compared with cows injected with PGF2α and lutectomized (Buford et al., 1996). Lemaster et al. (1999) tested the possibility of a role for oxytocin. Administration of oxytocin on day 5 through 8 decreased the embryonic survival in progestogen-supplemented beef (pregnancy rate: 33.3%) even when it was accompanied by lutectomy (30%). However, pregnancy rate was restored to normal (80%) if oxytocin-treated cows also received a prostaglandin synthesis inhibitor. Therefore, the oxytocin acted through stimulation of uterine secretion of PGF2α. Given that a regressing CL was required to manifest the negative effect of PGF2α on embryo survival (Buford et al., 1996; Seals et al., 1998), it could be concluded removal of luteal tissue reduced the lethal effect by interruption of the modulatory effect of oxytocin on uterine PGF2α secretion (Lemaster et al., 1999) and by abolition of PGF2α secreted from CL once luteolysis was initiated (Rexroad and Guthrie, 1979). Therefore, a direct embryotoxic effect of oxytocin on embryo development is improbable. Niswender et al. (2007) summarized evidence that luteal PGF2α is involved in luteolysis in sheep. Once the uterine signal (PGF2α) initiates increased luteal secretion of PGF2α, it triggers mobilization of Ca ++ in large luteal cells and secretion of oxytocin by these cells. Oxytocin in turn, triggers mobilization of Ca ++ in small luteal cells. Apoptosis follows Ca ++ mobilization in both cell types. Thus, secretions from the CL are involved in its demise, acting within the CL itself. As conclusion, [Figure - 5] indicates the sequence of endocrine events conducive to embryonic death or survival in short and normal estrous cycles. [68] References
Copyright 2008 - Archivos Latinoamericanos de Produccion Animal The following images related to this document are available:Photo images[la08023t4.jpg] [la08023t2.jpg] [la08023f4.jpg] [la08023t3.jpg] [la08023f1.jpg] [la08023f5.jpg] [la08023f3.jpg] [la08023f2.jpg] [la08023t1.jpg] |
| |||||||||

![[Figure - 1]](/showimage?la/photo/la08023f1.jpg){kind=link}
![[Figure - 2]](/showimage?la/photo/la08023f2.jpg){kind=link}
![[Figure - 3]](/showimage?la/photo/la08023f3.jpg){kind=link}
{kind=link}
![[Figure - 4]](/showimage?la/photo/la08023f4.jpg){kind=link}
![[Figure - 5]](/showimage?la/photo/la08023f5.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}