 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Archivos Latinoamericanos de Produccion Animal, Vol. 16, No. 4, October-December, 2008, pp. 224-233 Original Article Comparación de modelos en la evaluación genética de caracteres de crecimiento del ganado Brahman en el sureste de México Comparison of models in genetic evaluation o. f growth traits for Brahman cattle in southeastern Mexico. RJ Estrada-León, JG Magaña Monforte, JC Segura Correa Facultad de Medicina Veterinaria y Zootecnia, Universidad Autónoma de Yucatán, Mexico Correspondence Address:J G Magaña Monforte, Facultad de Medicina Veterinaria y Zootecnia, Universidad Autónoma de Yucatán. Km 15.5 Carretera Mérida- Xmatkuil, AP 4-116, Mérida, Yucatán, Mexico, jmagana@uady.mx Date of Submission: 07-Feb-2008 Code Number: la08031 Abstract Variance components and genetic parameters for birth weight, BW (n=2905), adjusted weaning weight to 205 days, WW205 (n=2264) and preweaning daily gain, ADG (n=2264) in four herds of Brahman cattle in southeastern Mexico were estimated. Six animal models were fitted for each trait, ranging from a simple model with the additive direct effect (Ó2a) as the only random effect to the complete model with Ó2a, additive maternal effect (Ó2m), covariance between them (óam ) and maternal permanent environmental effect (Ó2c). The six models included as a fixed effects contemporary group of birth (Herd-Year-Season), Sex and parity number. Likelihood ratio test, prove for all traits, Ó2m and óam were important (P<0.05) but not Ó2c (P>0.05). Ignoring maternal effects in the models tended to increase direct heritability and also, ignoring óam tended to increase error. Subtract Ó2c not affect direct and maternal heritability. Spearman correlations of direct and also for maternal animal genetic values, between models with and without Ó2c were 1, suggesting no differences in the ranking of animal. The genetic correlations between additive and maternal heritabilities for all characteristics were negative, affecting estimated total heritability (Lower than in models omitting óam). The additive genetic variance given maternal effects (Ó2a.m) was constant across models suggesting a correct partition of additive genetic effects into their direct and maternal components. In conclusion, the most appropriate model must be selected for each characteristic, in order to reduce bias in genetic evaluation.Keywords: Growth traits, Models, Genetic Parameters, Breeding Values, Brahman, Tropic RESUMEN Componentes de (Co) varianza y parámetros genéticos para Peso al Nacimiento, PN (n=2905), Peso Ajustado a 205 días, PA205 (n=2264) y Ganancias de Peso Predestete, GDP (n=2264) fueron estimados en cuatro hatos de ganado Brahman en el Sureste de México. Seis modelos animal fueron ajustados, desde el más simple con únicamente los efectos aditivos directos (Ó2 a ), hasta el completo que incluyó Ó2 a , los efectos aditivos maternos (Ó2 m ), la covarianza entre ambos (Óam ) y los efectos ambientales permanentes de la madre (Ó2 c ). Los seis modelos incluyeron como efectos fijos el grupo contemporáneo de nacimiento (hato-año-época), el sexo y el número de parto. La prueba de razón de verosimilitudes demostró que para todas las características evaluadas,Ó2 m y óam fueron importantes (P<0.05). Ignorar ó2 m tiende a incrementar la indice de herencia directa, así como también, ignorar óam tiende a incrementar los errores. La eliminación de Ó2 c, no afectó las estimaciones. Las correlaciones de Spearman para los valores genéticos directos y maternos de los animales entre los modelos con y sin Ó2 c fueron de 1, lo que sugirió no diferencia en la jerarquización de los animales. Las correlaciones genéticas entre las indice de herenciaes directas y maternas para todas las características fueron negativas, lo que afectó la estimación de la indice de herencia total (menor que en modelos omitiendo óam). La Ó2 a dado los efectos maternos (Ó2 a.m ) fue constante, sugiriendo una correcta partición de los componentes directos y mater nos. En conclusión, se debe seleccionar el modelo apropiado para cada característica, con la finalidad de reducir el sesgo en las evaluaciones genéticas. Palabras clave: caracteres de crecimiento, modelos, parámetros genéticos, valores genéticos, Brahman, Trópico. Introducción Los programas actuales de mejora genética para el ganado de carne, deben enfocarse hacia caracteres de importancia económica, como crecimiento de be-cerros, calidad de la canal y rendimiento materno, entre otros (Diaz y Quintanilla, 2002). Los caracteres de crecimiento, se encuentran influenciados no solo por el efecto de los genes here-dados, sino también por efectos maternos, los cuales son considerados como efectos ambientales para el becerro y están determinados por factores genéticos y ambientales de la madre (Koch, 1972; Willham, 1972, 1980; Beker, 1980). La estimación de compo-nentes de (co)varianza para éstos caracteres, requie-ren de modelos apropiados (Willham, 1972); los mé-todos antiguos dependían de diferentes tipos de covarianza en el parentesco de animales (Thompson, 1976; Cantet et al., 1988), pero en la actualidad, se han podido introducir modelos más realistas como "el modelo animal con efectos maternos", el cual ha mejorado la exactitud (Meyer, 1997; Bidanel, 1998). Sin embargo, esto no garantiza la confiabilidad de las estimaciones (Naal y Segura-Correa, 2004). Una de las principales fuentes de error, es la elec-ción del modelo genético a utilizar en el análisis de los datos, (Meyer, 1997; Clement et al., 2001). Como consecuencia, puede modificarse la jerarquización de los animales y generar una reducción en la res-puesta esperada a la selección. El presente trabajo tiene como objetivo identificar el modelo apropiado para caracteres de crecimiento, determinar el efecto de otros modelos sobre la estima-ción de los parámetros genéticos y la jerarquización de los valores genéticos del ganado Brahman en el Sureste de México. Materiales y Métodos Origen de la información y manejo de los animales. El presente trabajo, se realizó en el Estado de Yucatán, México, localizado entre los paralelos 19º40′ y 21º37′ de latitud norte y los meridianos 87º32′ de longitud oeste (INEGI, 2004). El clima de la región es tropical subhúmedo con lluvias abundantes en vera-no (Aw0), con promedios de temperatura mensual y precipitación pluvial al año de 25,8°C y 1105 mm, respectivamente. Los hatos estaban afiliados a la Asociación Mexicana de Criadores de Cebú (AMCC), contaban con praderas establecidas de pasto Guinea (Panicum maximum) y áreas de pasto de corte Taiwan (Penicetum purpurium) y se dedicaban a la produc-ción de sementales y hembras de pie de cría de la raza Brahman, que se utilizan en varios hatos distri-buidos tanto en la región del trópico de México como en algunos países de Centroamérica. Los animales se pesaron al nacimiento, se identi-ficaron con un tatuaje en la oreja para mantener la identificación de la madre y del padre. A partir de los dos meses de edad, recibieron un tratamiento contra parásitos internos y externos, así como las vacunas contra las enfermedades de mayor incidencia en la región durante la fase predestete, cuya duración fue de 8 meses aproximadamente. Al destete, se pesaron y marcaron con hierro candente utilizando un nú-mero de identidad. Los animales permanecieron en un sistema de pastoreo rotacional y durante la época seca del año (febrero a mayo) se suplementaron con pasto de corte, pollinaza y melaza y/o alimento ba-lanceado con 13% PC y 2.2 - 2.4 Mcal de EM /kg MS, a razón de 0.250 kg por animal al día. La reproduc-ción en los hatos se realizó través de inseminación artificial y monta controlada; la importación de se-men y sementales de los Estados Unidos, era una práctica común en los hatos evaluados. Estructura y edición de los datos Se recopilaron y capturaron los pesos al nacimien-to (PN) y pesos al destete (PD), así como también toda la información disponible de becerros Brahman regis-trados o no en la AMCC. A partir de ésta información, se calcularon las ganancias de peso predestete (GDP) y los PD se ajustaron a 205 días (PA205) (BIF, 2003). Datos extremos no lógicos y de más o menos tres des-viaciones típicas de la media fueron eliminados. Las identificaciones originales de los animales fueron codificadas acorde a su fecha de nacimiento mediante el programa Demograf (Carolino y Gamma, 2002). La conectividad de los grupos contemporá-neos se estimó utilizando el programa MILC (Fries, 1998), eliminado los no conectados. Después de la edición de los datos, se completó un archivo de pedigrí de 4531 animales, de los cuales 2905 contaban con PN y 2264 con PA205 Y GDP. Asimismo, para PN, se identificaron 112 padres y 962 madres de los animales con datos (2905); de éstos, 13 toros y 103 (10,7%) vacas tuvieron datos propios como becerros. Los padres fueron hijos de 9 toros (abuelos paternos) con un promedio de 48,7 nietos de ambos sexo. Las vacas (hijas de 42 toros), tuvieron un prome-dio 3.02 crías por vaca (el 47,9% de las 962 vacas tu-vieron 3 o más partos). Para PA205 y GDP, se identifi-caron 112 sementales y 761 madres de los animales con datos (2264); de éstos, 10 toros hijos de 6 sementales (abuelos paternos) con un promedio de 14.6 nietos y 82 (10,8%) vacas tuvieron datos propios como bece-rros. También, las vacas fueron hijas de 36 sementales (abuelos maternos) y en promedio tuvieron 2,98 crías por vaca (el 47,2% de las 761 vacas tuvieron 3 o más partos). La estructura de datos, cumple con las reco-mendaciones mínimas de 10% de madres con datos propios como becerras y más de 1,5 crías por vaca, para la correcta partición de los efectos maternos y ambientales permanentes (Maniatis y Pollot (2003). Estructuras similares han sido consideradas como adecuadas para la evaluación genética del peso al destete por Diaz et al. (2002) y por Cinachi et al. (2006), en ganado Limousine Español y en ganado Caracu Brasileño, respectivamente. Análisis estadísticos de los datos Los componentes de (Co) varianza y parámetros genéticos, fueron estimados mediante el procedimien-to de Máxima Verosimilitud Restringida (REML), ajustando un modelo animal univariado para PN, PA205 y GDP, utilizando el programa MTDFREML (Boldman, 1995). Toda la información disponible del pedigrí fue incluida en el análisis con la finalidad de minimizar sesgos e incrementar la precisión de la evaluación al adicionar conexiones entre los anima-les de cada hato (Meyer, 1997). Un análisis previo a través del procedimiento MIXED del paquete estadístico SAS (2002), fue reali-zado para determinar la inclusión de los efectos fijos; de acuerdo al resultado obtenido (P<0.05), se incluyó el efecto fijo de grupo contemporáneo (Hato- Año-Época de nacimiento) donde se consideraron 4 ha-tos, dos épocas (Lluvias y no lluvias) y 9 años (1995-2003), grupos contemporáneos con menos de tres observaciones fueron eliminados; también, se inclu-yeron los efectos fijos de sexo del becerro (machos y hembras) y el numero de parto de la madre (1-10). Para evaluar la inclusión de cada efecto aleatorio y seleccionar el mejor modelo para cada variable, se ajustaron los siguientes: El modelo 1 (D), incluyó únicamente el efecto aditivo directo; el modelo 2 (in-cluyó el efecto aditivo directo y materno con (Ó 2 ) = 0; el modelo 3 (DC) incluyó el efecto aditivo directo y el efecto ambiental permanente; el modelo 4 (DMR) in- cluyó los mismos efectos que el modelo 2 pero con (Ó 2 ) ≠ 0; el modelo 5 (DMC) incluyó adicionalmente al modelo 2 el efecto ambiental permanente con (Ó 2 ) = 0, y el modelo 6 (DMCR) similar al modelo anterior pero con (Ó 2 ) ≠ 0. Los modelos en notación matricial fueron: Modelo 1 (D) y = Xβ + Z 1a + e Modelo 2 (DM) y = Xβ + Z 1a + Z2m + e [Covarianza (a,m) = 0] Modelo 3 (DC) y = Xβ + Z1a + Z3c + e Modelo 4 (DMR)y = Xβ + Z1a + Z2m + e [Covarianza (a,m) = Aóam ] Modelo 5 (DMC)y = Xβ + Z1a + Z2m + Z3c + e [Covarianza (a,m) = 0] Modelo 6 (DMRC) y = Xβ + Z1a + Z2m + Z3c + e [Covarianza (a,m) = Aóam ] Donde: y = Vector de la variable de respuesta (PN, PA205, GDP), β es el vector que contiene los efectos fijos aso-ciado a la matriz de incidencia X, a es el vector que contiene los efectos genéticos aditivos directos aso-ciado a la matriz de incidencia Z1, m es el vector que contiene los efectos genéticos aditivos maternos de cada animal asociado a la matriz de incidencia Z2, c es el vector de efectos aleatorios del ambiente perma-nente de la vaca asociado a la matriz de incidencia Z3 y e es el vector de efectos residuales. Los supuestos estadísticos del modelo completo (DMRC) fueron:
Donde: A es la matriz de parentesco, Ó2a es la varianza del efecto genético aditivo directo, Ó2m es la varianza del efecto aditivo materno, ó2c es la covarianza entre los efectos genéticos aditivo directo y materno, es la varianza del efecto ambiental perma-nente del efecto materno, Ó2e es la varianza del error, V es número de vacas, n es el número de observacio-nes e I es la matriz identidad. El criterio de convergencia basado en la función verosimilitud (-2log L) fue de 10 -9. Se realizaron va-rios análisis tomando como valores iníciales los re-sultados del análisis anterior y se consideró que la convergencia había sido alcanzada cuando la varia-ción en los valores de la función verosimilitud fue menor que 10 -6 . La comparación estadística entre pares de modelos a evaluar para determinar la inclusión de un efec-to aleatorio, se realizó mediante la prueba de razón de verosimilitudes (PRV), que determina la diferen-cia entre los logaritmos de las funciones de verosimi-litud (-2log L) estimadas y las compara con un valor de Chi cuadrada (χ2). La PRV se representa como: λ= -2(Log L del modelo completo [b] - Log L del modelo reducido [a]) Donde: Log L= Logaritmo de verosimilitud, a= modelo con n parámetros y b= modelo con n+1 parámetros. Si λ ≥ χ21,α 0.95, el modelo [b] se acepta significativamente como de mejor ajuste que el modelo [a], de lo contrario no son diferentes estadísticamente (Lynch and Walsh, 1998). La indice de herencia total (h2t) fue estimada como: h2t = (Ó2 a + 1.5Óam + 0.5Ó2 m) / Ó2 p; donde Ó2 p = Ó2 a + Ó2 m + Ó2 am + Ó2 c+ Ó2 e También, con la finalidad de encontrar eviden-cias de problemas al intentar partir los efectos genéticos en directos y maternos, se estimó la varianza genética aditiva dados los efectos maternos (Ó2a.m) y la indice de herencia directa condicional (h 2a.m) (Meyer, 1992) como: Ó2 am = Ó2 a - Ó2 am / Ó2 m ; h 2a.m = Ó2 am/Ó2 p Asimismo, con la finalidad de analizar los cam-bios en la jerarquización de los animales a través de los modelos evaluados, se estimaron los coeficientes de correlación de rangos de Spearman entre los valo-res genéticos directos y maternos de los animales es-timados para cada modelo evaluado, utilizando el paquete estadístico SAS (2002). Resultados y Discusión El valor promedio de PN, fue de 33.10 ±4.65 kg., el cual fue ligeramente mayor al reportado para gana-do Brahman por Plasse et al. (2002b), Pico et al., (2004) y por Parra-Bracamonte et al. (2007) en Venezuela, Sudáfrica y México respectivamente, que van de 30.8 a 32.3 kg. Con respecto a PA205 y GDP los valores promedio fueron de 204.05±37.71 kg, y 0.840±0.18 kg., respectivamente. El valor de PA205, se encuentra dentro del rango reportado por los mismos autores, que van de 175 a 212 kg. Los resultados de las pruebas de razón de verosi-militudes, los componentes de (co)varianza y los parámetros genéticos para PN, PA205 y GDP, a tra-vés de los diferentes modelos evaluados se presen-tan en los [Cuadro - 1], [Cuadro - 2] y [Cuadro - 3] respectivamente. Peso al Nacimiento (PN) Para PN, el modelo D, produjo un valor para la varianza genética directa (Ó2a) alto de 9.89 kg 2 y conse- cuentemente para la indice de herencia directa (h2d) de 0.52; la adición del efecto ambiental permanente (c2) (modelo DC), mostró un mejor ajuste, identificando un aporte de los efectos de la madre (vaca) del 2.7%, sin embargo éste no fue importante (P>0.05). Por el con-trario, cuando el efecto aditivo maternos (h2m) fue in-cluido en el modelo D (modelo DM), en lugar del efecto ambiental permanente, se produjo un mayor ajuste, ya que la inclusión de éste, si fue estadísticamente signi-ficativo, estimándose una indice de herencia materna del 5% y una indice de herencia directa de 0.42, la cual se redujo en un 23.8% en comparación con la estima-da con el primer modelo D (0.52). La influencia de los efectos maternos sobre PN, son considerados de baja magnitud y se deben a efec-tos citoplásmicos como los sugeridos por Tess y Robison (1990) y de medio ambiente uterino provisto por la vaca durante la gestación, (Maurer y Gregory, 1990). Autores como Meyer (1992), Waldron et al., (1993), Ferreira et al. (1999), Dominguez-Viveros et al. (2003), Parra (2006), entre otros, han mostrado la im-portancia de los efectos maternos para ésta carac-terística en bovinos de carne de diferentes razas, a través de comparación de modelos, observándose una sobre estimación de la varianza aditiva directa cuan-do los efectos maternos existen y son omitidos. La adición de h2m al modelo DC (modelo DMC), mostró de manera similar un mejor ajuste (P<0.05), mostrando que para PN no fue importante c2, ya que su valor fue 0.0. La prueba de razón de verosimilitud (PRV) demostró que la adición del efecto ambiental permanente no mejoró (P>0.05) ninguno de los mo-delos que la incluyó, aceptando la hipótesis de igual-dad a cero cuando se compararon los modelos DC contra D; DMC contra DM y DMRC contra DMR [Cua-dro 1], por lo que ignorar éste efecto no causa modifi-caciones sobre el ajuste del modelo y tampoco sobre las estimaciones de h2d y de h2m. Resultados similares sobre la no significancia de éste efecto, han sido re-portados por Domiguez-Viveros et al. (2003) y por Parra (2006) en ganado Tropicarne y Brahman de registro en México, sin embargo, en algunos casos aunque la magnitud sea pequeña, ésta debe incluirse al modelo si su eliminación tiende a incrementar los efectos aditivos maternos, inflando h2m (Meyer, 1992), lo que no sucedió en la presente evaluación. Por el contrario, la adición de la covarianza entre los efectos aditivos directos y maternos (óam ) a los modelos DM y DMC, mejoró de manera importante el logaritmo de verosimilitud (P<0.05), demostrando a través de PRV que se rechaza la hipótesis nula (Esti-mación = 0.00) al comparar los modelos DMR contra DM y DMRC contra DMC. A través del ajuste de es-tos modelos, se observó un efecto antagónico entre los efectos directos y maternos, estimándose una co-rrelación de -0.55, con importantes ajustes en las esti-maciones, ya que h2d y h2m , se incrementaron y el error disminuyó un 8%. Efectos antagónicos y de impor-tancia para PN han sido reportados por Meyer (1992), Ferreira et al., (1999), Domiguez-Viveros et al., (2003), Parra (2006), para bovinos de carne con diferentes magnitudes, lo que difiere de Waldron et al., (1993) quien ha reportado estimaciones positivas. La varianza genética aditiva dados los efectos maternos (Ó2a.m), no mostró variaciones importantes través de los modelos evaluados para PN, lo que su-giere una correcta partición entre los efectos directos y maternos para ésta característica [Cuadro - 2]. Por lo tanto el mejor modelo para PN en la presen-te evaluación fue DMR. Los resultados coinciden con lo estimado a través de las correlaciones de Spearman, las cuales fueron de 1 entre los valores genéticos di-rectos y maternos de los animales, estimados a través del modelo completo (DMRC) y el modelo elegido (DMR) [Cuadro - 4], sugiriendo no cambios en la jerarquización de los animales [Cuadro - 4] La indice de herencia total estimada fue de 0.40, debido al antagonismo entre los efectos directos y maternos, observándose a través de los diferentes modelo que éste valor puede sobre estimarse de 4 a 12 unidades, así mismo, la indice de herencia directa condicional (h2a.m) estimada fue de 0.39 [Cuadro - 3]. Peso al Destete Ajustado a 205 días (PA205). Para PA205, el modelo D, produjo un valor alto para Ó2 (483.63 kg 2 ), estimándose una h2d de 0.62; la adición del efecto c2 (modelo DC), mostró un mejor ajuste, identificando un aporte importante (P>0.05) de los efectos de la vaca de un 21%. Así mismo, cuan-do el efecto aditivo maternos (h2m) fue incluido al modelo D (modelo DM), en lugar del efecto ambiental permanente, se produjo un mayor ajuste (P<0.05) de un 3%, estimándose una h2m de 0.24 y una h2d de 0.31, misma que se redujo un 50%, en comparación con la estimada en el modelo D (0.62). La importancia de los efectos maternos para peso al destete, han sido ampliamente documentados (Koch, 1972; Willham, 1972, 1980; Beker, 1980); identificándose que para caracteres de crecimiento postnatales, la producción de leche es el principal efecto materno, estimándose una correlación entre ambos de 0.8, aunque también influyen los cuidados que le proporcione la vaca a su becerro (habilidad materna) (Meyer et al., 1994). La adición de h2m al modelo DC (modelo DMC), mostró de manera similar un mejor ajuste que los mo-delos D y DC, mostrando que c 2 no fue importante para PA205, ya que su valor fue 0.0004 y su inclusión no modificó las indice de herenciaes estimadas en el mo-delo DM. PRV, demostró que la adición del efecto am-biental permanente no mejoró estadísticamente nin-gún modelo que la incluyó, aceptando la hipótesis de igualdad a cero cuando se compararon los modelos DC contra D; DMC contra DM y DMRC contra DMR [Cuadro - 1], por lo que ignorar éste efecto no causa mo-dificaciones sobre el ajuste del modelo y tampoco so-bre las estimaciones de h2d y de h2m. Resultados simila-res sobre la inclusión del efecto ambiental permanente (c) para pesos ajustados a 205 días y peso ajustado a 8 meses de edad, han sido reportados por Meyer (1992) para cruzas de cebú en Australia y por Domínguez-Viveros, et al. (2003) para ganado Tropicarne en Méxi-co respectivamente. Por el contrario, Robinson (1996) reportó para ganado Angus Australiano, que la inclu-sión del efecto ambiental permanente fue importante para los modelos DM y DMR. De manera similar, otros autores han decidido incluir en sus modelos genéticos al efecto ambiental permanente para evaluar pesos ajustados a 205 días de ganado Brahman, debido su magnitud en los resultados y a la naturaleza de la característica, la cual incluye mediciones repetidas por vaca (Plasse, et al., 2002a; Plasse et al., 2002b; Plasse, et al., 2004; Pico, et al., 2005), sobre todo si modifican las estimaciones de h2m. Asimismo, de acuerdo a PRV, la adición de óam a los modelos DM y DMC, mejoró de manera impor-tante el logaritmo de verosimilitud (P<0.05), por lo que se rechaza la hipótesis nula (Estimación = 0.00) cuando se compararon los modelos DMR contra DM y DMRC contra DMC. A través del ajuste de estos modelos, se observó un efecto antagónico entre los efectos aditivos directos y los maternos, estimándose una correlación de -0.46, con importantes ajustes, ya que el error disminuyó un 5% y las estimaciones de h2d y h2m, se incrementaron un 11% y 13% respectiva-mente. Resultados similares aunque de mayor mag-nitud han sido reportados por Pimenta et al., (2001) (-.68) en ganado Guzerat y por Meyer (1994) -.0.74 y - 0.78 para DMR y DMRC respectivamente y de menor magnitud por Domínguez-Viveros et al. (2003) de - .36 y -.37 para DMR y DMCR respectivamente. Valores muy elevados de óam, han sido reportados por la literatura, justificándose su estimación de una manera escéptica; sin embargo Cundiff (1972) men-ciona que los efectos adversos débiles entre los efec-tos directos y maternos son considerados más reales y creíbles. Dentro de los factores que podrían incre- mentar la óam , se encuentra una variación extra no considerada, como puede ser grupos de manejo pri-vilegiados que incrementen la covarianza entre hijos de un semental dentro del grupo contemporáneo, o una variación mayor entre sementales que entre hem-bras debida a una mayor varianza genética por la importación de sementales no relacionados de alto valor genético (Robinson, 1996; Meyer, 1997), situa-ción que ocurre en los hatos evaluados. Otro de los factores, pudiera ser la falta de conectividad de los sementales, que de manera simi-lar podría estar aportando una variación extra a tra-vés de los años de evaluación, creando una interacción sementales x años o sementales x grupos contemporáneos. El problema de conectividad entre los grupos contemporáneos en las evaluaciones genéticas está relacionado con la eliminación de los "parámetros ruidosos" como las interacciones debi-do a la estimación de un efecto genético de grupo (Foulley et al., 1990), sin embargo en la presente eva-luación, la conectividad fue verificada. Al respecto diversos autores han evaluado mode-los alternativos que incluyan el efecto de la interacción semental x año o por grupo contemporá-neo como un efecto aleatorio adicional, encontrando que la adición de éste factor reduce drásticamente el -2log y se reporta como estadísticamente significati-va su inclusión al modelo, disminuyendo la magni-tud de la correlación negativa entre los efectos direc-tos y maternos, haciendo más confiables las estima-ciones (Robinson, 1996; Meyer, 1997; Lee y Pollak, 1997; Pico et al., 2004). Sin embargo, esto no pudo realizarse en la presente evaluación debido a la es-tructura de datos y reducida información. La varianza genética aditiva dados los efectos maternos (Ó2a.m), no mostró variaciones importantes través de los modelos evaluados para PA205, sugi-riendo una correcta partición entre los efectos direc-tos y maternos [Cuadro - 2]. Por lo tanto el mejor modelo para PA205 en la pre-sente evaluación fue DMR. Los resultados coinciden con las correlaciones de Spearman estimadas entre los valores genéticos directos y maternos con el mo-delo completo (DMRC) y el modelo elegido (DMR), las cuales fueron de 1, lo que sugiere no cambios en la jerarquización de los animales [Cuadro - 4]. La indice de herencia total estimada fue de 0.33, debi-do a un antagonismo entre los efectos aditivos directos y maternos, observándose a través de los diferentes mode- los que éste valor puede sobre estimarse de 2 a 29 unida-des, así mismo, indice de herencia directa condicional (h2a.m) estimada fue de manera similar 0.33 [Cuadro - 3]. Ganancias de Peso Predestete (GDP). GDP, siguió mostrando la misma tendencia que PN y PA205, a través del modelo D, produjo el mayor valor para Ó2a , que en los demás modelos ajustados, estimándose una h2d de 0.54; la adición de c2 (modelo DC), mostró un mejor ajuste, identificando un aporte importante (P<0.05) de los efectos de la vaca de un 19%. También, cuando el efecto aditivo materno (h2m) fue incluido en el modelo D (modelo DM), se produjo un mejor ajuste que en DC de un 2%, estimándose una h2m de 0.21 y una h2d de 0.24, misma que se redujo en 30 unidades en comparación con la estimada con el pri-mer modelo D (0.54). La adición de h2m al modelo DC (modelo DMC), mostró un mejor ajuste que los mode-los D y DC, mostrando que c 2 no fue importante para PA205 y su inclusión no modificó las indice de herenciaes estimadas en el modelo DM. PRV, demos-tró que la adición de c 2, no mejoró estadísticamente ninguno de los modelos que la incluyó, aceptando la hipótesis de igualdad a cero cuando se compararon los modelos DMC contra DM y DMRC contra DMR, por lo que ignorar éste efecto no causa modificaciones sobre el ajuste del modelo y tampoco sobre las estima-ciones de h2a y de h2m. En contra parte, de acuerdo a PRV, la adición de Ó 2 a los modelos DM y DMC, me-joró de manera importante el logaritmo de verosimili-tud (P<0.05), por lo que se rechaza la hipótesis nula (Estimación = 0.00) cuando se compararon los mode-los DMR contra DM y DMRC contra DMC. A través del ajuste de estos modelos, se observó un efecto anta-gónico entre los efectos aditivos directos y los mater-nos, estimándose una correlación de -0.42, con impor-tantes ajustes, ya que el error disminuyó un 5% y las estimaciones de h2d y h2m, se incrementaron un 11% y 13% respectivamente. La varianza genética aditiva dados los efectos maternos (Ó2a.m), no mostró variaciones importantes través de los modelos evaluados, sugiriendo una co-rrecta partición entre los efectos directos y maternos en para GDP en la presente evaluación [Cuadro - 2]. Por lo tanto el mejor modelo para GDP en la pre-sente evaluación fue DMR. Los resultados coinciden con lo estimado a través de las correlaciones de Spearman, ya que a través de los valores genéticos directos y maternos estimados entre el modelo com-pleto (DMRC) y el modelo elegido (DMR) se obtuvo un valor de 1 [Cuadro - 4], lo que sugiere no cambios en la jerarquización de los animales [Cuadro - 4]. La indice de herencia total estimada fue de 0.28, debido a un antagonismo entre los efectos aditivos directos y maternos, observándose a través de los di-ferentes modelo que éste valor puede sobre estimarse de 1 a 26 unidades, así mismo, la indice de herencia directa condicional (h2a.m ) estimada fue de 0.26 [Cuadro - 3]. Conclusión Para las características de crecimiento predestete del ganado Brahaman en la presente evaluación, el mejor modelo fue DMR, el cual incluyó los efectos aditivos directos, los maternos y la covarianza entre ambos, ya que se demostró que los efectos ambienta-les permanentes no fueron importantes; la estructura de datos y estructuras familiares, juegan un papel importante para la correcta partición de la varianza fenotípica en sus compontes, lo que en la presente evaluación pudo analizarse a través de la varianza genética cuando los efectos maternos están presen-tes, en diferentes modelos, así como también es im-portante estimar la indice de herencia total y la indice de herencia condicionada, ya que se consideran más precisas. Así mismo, la elección de un modelo inade-cuado para el análisis de éstas características tiene repercusiones sobre las estimaciones de los compo-nentes de varianza y por consiguiente, sobre los parámetros genéticos y los valores genéticos (Dife-rencia Esperada de la Progenie DEP′s) de los anima-les, lo que influye sobre la jerarquización de los mis-mos y podría afectar la respuesta esperada a la selec-ción. La elección de un modelo adecuado reduce sesgos en la evaluación genética. Agradecimiento A la Fundación Produce Yucatán AC por el financiamiento del presente trabajo y el primer autor agradece al Consejo Nacional de Ciencia y Tecnolo-gía de México (CONACYT) por la beca para realizar estudios de posgrado. A la Asociación Regional de Criadores de Ganado Brahman de Registro por las facilidades otorgadas para el presente estudio. Final- mente a los Dres. Miguel A. Toro I., Clara Diaz M., Ma. Jesus Carabaño y Jesús Fernández M., del De-partamento de Mejora Genética del Instituto Nacio-nal de Tecnología Agraria y Alimentaria (INIA), Ma-drid, España, por sus valiosas recomendación y apo-yo durante la realización del presente trabajo. [39] References
Copyright 2008 - Archivos Latinoamericanos de Produccion Animal The following images related to this document are available:Photo images[la08031t2.jpg] [la08031t1.jpg] [la08031t4.jpg] [la08031t3.jpg] |
| |||||||||

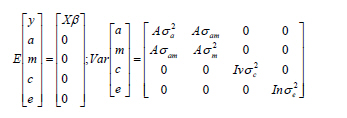
![[Cuadro - 1]](/showimage?la/photo/la08031t1.jpg){kind=link}
![[Cuadro - 2]](/showimage?la/photo/la08031t2.jpg){kind=link}
![[Cuadro - 3]](/showimage?la/photo/la08031t3.jpg){kind=link}
![[Cuadro - 4]](/showimage?la/photo/la08031t4.jpg){kind=link}