 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
East African Journal of Public Heath, Vol. 2, No. 2, Oct, 2005, pp. 28-31 Emergence of Mutations Associated with Resistance to Sulfadoxine-Pyrimethamine (SP) After Single Therapeutic Dose: Implications on the Useful Therapeutic Life of SP in Malaria Holoendemic Areas D.S. Tarimo Depat. of Medical Parasitology &
Entomology, School of Public Health & Social Sciences Code Number: lp05015 Abstract Background: Drugs with long elimination
half-lives such as Sulfadoxine –Pyrimethamine (SP) maintain sub-curative levels
in blood for a long time such that in high malaria transmission areas,
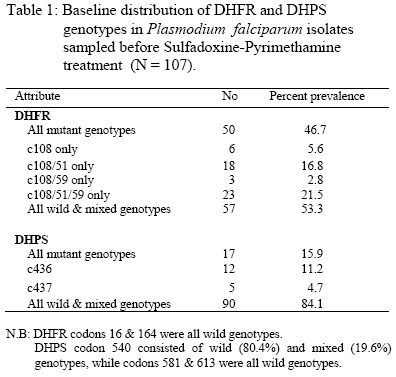
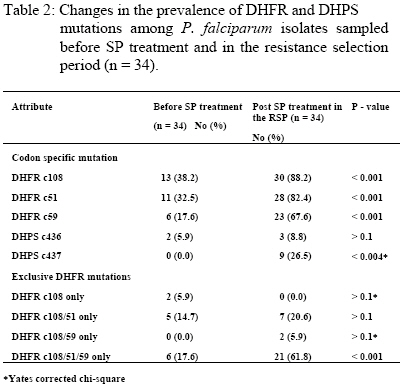
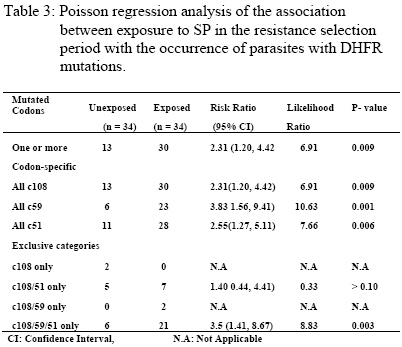
re-infecting parasites are continuously under selection pressure for resistant genotypes. Key words: Plasmodium falciparum, genetic resistance, holoendemicity, Sulfadoxine-Pyrimethamine. Introduction Drugs with long elimination half-life such as SP have high propensity to selection for resistant (mutant) genotypes at re-infection because of exposure to sub-curative drug levels (1). Selection for P.falciparum mutant genotypes occurs in the genes encoding for the parasite dihydrofolate reductase (DHFR) and dihydropteroate synthetase (DHPS) enzymes (2,3) targeted by Pyrimethamine and Sulfadoxine respectively (4). DHFR mutation Ser→Asn 108 causes moderate Pyrimethamine resistance in-vitro, with high level resistance resulting from the addition of Asn→ Ile 51 and/or Cys→ Arg 59 mutations; while DHFR mutation Ile→ Leucine 164 in combination with Ser→ Asn 108 and Cys→ Arg 59 and/or Asn→Ile 51 is associated with very high level of resistance to Pyrimethamine (5). DHPS mutations that have been associated with in-vitro resistance to Sulfadoxine include Ala®Gly 437 that seems to be especially important for resistance to Sulfadoxine, while additional mutations Ser→ Phe 436, Ser→ Ala 436, Lys→ Glu 540, Ala→ Gly 581 and Ala→ Thr/Ser 613 mediate high level resistance to Sulfadoxine (6). The polymerase chain reaction (PCR) and restriction digestion provides a useful molecular tool for the detection of point mutations in the genes encoding for DHFR and DHPS enzymes (7, 8). Mutations in the DHFR & DHPS enzymes represent low-grade resistance that cannot be detected by the currently recommended in vivo methods for assessing P.falciparum resistance to antifolates in high transmission areas (9), but can only be detected in vivo by molecular assays (7). Reported is the prevalence of DHFR and DHPS mutations in P.falciparum isolates sampled before SP treatment and at re-infection in a holoendemic area of Tanzania. The aim was to test the hypothesis that, among re-infections occurring in the resistance selection period (RSP), parasites with mutations (resistance) will survive in the presence of sub-curative levels of SP whereas parasites with wild / mixed genotypes (sensitive) will be cleared (10), and assuming no mutations arise de-novo spontaneously or mutagenically (11),in 3 – 4 asexual cycles after infection, surviving (resistant / mutated) parasites would build up to reach microscopic patency and therefore parasites sampled in the RSP (1) will have a higher prevalence of mutations than parasites sampled before SP treatment to indicate selection by SP. This study was approved by the Tanzanian Ministry of Health, and consent for participation obtained from mothers or guardians on behalf of the children. Patients and methods Enrollment and follow up The study was part of a larger study that examined the clinical efficacy of CQ and SP carried out in the holoendemic Kibaha district, coastal Tanzania prior to policy change to SP. Children 12 – 59 months old, assigned to receive SP had their parasites sampled before treatment with standard doses of oral SP (Fansidar® F.Hoffman – LaRoche, Basel, Switzerland: 25mg Pyrimethamine & 500 mg Sulfadoxine). Parasitaemia was normally assessed every after 24 hours, and at any other time as the clinical condition of the child dictated. At enrollment and thereafter, a blood film was pronounced negative when the examination of 100 fields was negative after 24 hours lapse from the last positive film (12). Treatment outcome was evaluated clinically and parasitologically as described (9,13). Blood samples for DHFR and DHPS genes assays were drawn pre-treatment on day 0, then day 7, 14, 21, 28 & 56 or any other day if the child was ill and parasitaemic. The extended period of follow up was chosen based on the long terminal elimination life of SP which results in sub-curative levels that maintain a resistance selection period extrapolated from pharmacokinetic data to last between days 15 – 52 after SP treatment (1). All blood samples were put into eppendorf tubes with EDTA and kept frozen (-400C) until use. DNA extraction and amplification Frozen blood samples were thawed, digested with proteinase K and incubated at 370C overnight. P.falciparum genomic DNA was extracted by chloroform/isoamyl alcohol – phenol and precipitated by ethanol to obtain a DNA pellet. The DNA was amplified by a PCR method described (8) with some modifications. An outer PCR was performed simultaneously for genotyping DHFR and DHPS using primers targeting both genes in the same reaction (primers M1 & M5 for DHFR and primers R1 & R2 for DHPS). The outer PCR produced fragments of 648 bp (DHFR) and 710 bp (DHPS) that were verified by running the positive controls on a 1.5% agarose gel. Four separate nested PCR reactions were then performed. For DHFR, bands of 522 bp and 326 bp were produced using primers M3&F/ and M4 & F/, respectively. For DHPS, bands of 438 bp and 161 bp were produced using primers K&K/ and L-&L/, respectively. The nested PCR products were digested by a set of restriction enzymes so as to genotype the specific codons. M3&F/PCR product was digested by Tsp5091 (codon 51), BsrI and AluI (codon 108), NIaIII (codon 16) and DraI (codon 164). M4&F PCR product was digested by XmnI (codon 59), BsrI and AluI (codon 108). K&K/ PCR product was digested by MnII & MspAI (codon 436), AvaII and MwoI (codon 437) and FokI (codon 540). L-&L/ PCR product was digested with BstUI and BsII (codon 581), MwoI and BsaWI (codon 613). All the enzymes were from New England Biolabs, Inc. The digest products were applied to a 2-2.5% Metaphore Agarose electrophoresis gel, stained with Ethidium Bromide and the bands observed in UV-light. Analysis The data was entered into SPSS 10.0 for Windows and analysed. Polymorphisms in the wild (sensitive), mutant (resistant) and mixed (wild / mutant) genotypes of DHFR (codons c108, c59, c51, c16 & c164) and DHPS (codons c436, c437, c540, c581 & c613) were assessed in parasites sampled before treatment and in the resistance selection period (RSP). To assess possible selection for mutations, the change in prevalence of codon-specific mutations and exclusive mutation categories was assessed among parasites sampled before SP treatment and in the RSP. The chi-square test for two-tailed significance was used to assess the difference in prevalence of mutations before SP treatment and during the RSP. A Poisson regression analysis was used to assess the association between exposure in the RSP span with the occurrence of one or more DHFR mutations, codon-specific mutations and exclusive mutation categories. Significance was set at the 0.05 level. Results A total of 119 children were enrolled, 107 completed follow up in the resistance selection period when 34 got malaria recurrence. The clinical (n = 110) and parasitological (n = 112) efficacy to SP was 98.2%, with only 1.8% treatment failures on day14 and 1.8% RII responses. P.falciparum genomic DNA were successfully extracted from 107 parasite isolates sampled before SP treatment and 34 parasite isolates sampled during the resistance selection period, and were successfully assayed for DHFR and DHPS genotypes. The genotypes were classified as wild (sensitive), mutant (resistant) and mixed (wild / mutant) types. The baseline distribution of DHFR and DHPS genotypes in Plasmodium falciparum isolates sampled before SP treatment (N = 107) is shown in Table 1. DHFR genotypes occurred as single, double or triple codon mutations among 46.7% of P.falciparum isolates sampled before treatment, of which the triple c108/51/59 and double c108/51 mutations were the most common. More than half of the pre-treatment infections (53.3%) thus carried wild or mixed DHFR genotypes. DHPS mutations in pre-treatment infections were very few (15.9%) and occurred only at codon c436 and c437. Majority of the pre-treatment infections (84.1%) thus carried wild or mixed DHPS genotypes. All infections carrying DHFR and DHPS mutant genotypes were subsequently cleared by the SP treatment. The prevalence of codon – specific mutations at DHFR codon c108, c51 and c59; and DHPS codon c437 was significantly higher among parasite isolates sampled in the terminal elimination phase of SP (the resistance selection period) than before treatment (Table 2). However, considering exclusive mutation categories, the prevalence of the triple DHFR mutation at codons c108/51/59 was significantly higher among parasites sampled in the terminal elimination phase of SP than before treatment. A Poisson regression analysis of the association between exposure to SP in the RSP with the occurrence of parasites with DHFR mutations (Table 3) showed that, the overall occurrence of DHFR mutations (one or more mutated codons) was strongly associated with parasites exposure to SP in the RSP. Mutations at specific codons were also strongly associated with exposure to SP. Of the exclusive mutation categories, only the triple c108/59/51 mutation was strongly associated with exposure to SP. Discussion The occurrence of a significantly high prevalence of DHFR c108, c59 & c51 codon – specific and exclusive c108/59/51 mutations, and DHPS c436 & c437 codon-specific mutations in parasites sampled in the RSP span indicates selection for these mutations. The strong association of exposure to SP in the RSP span with the occurrence of parasites with DHFR mutations provides an in vivo molecular basis for the earlier findings that, in vitro, parasites sampled in the RSP span are more likely to have resistant than sensitive genotypes (1,14). In the RSP span, SP concentration is certainly < Cmin but when parasites population consisting of sensitive and resistant (mutated) strains are exposed, the sensitive strains are cleared leaving the resistant strains to build up and manifest clinically (10,11) as observed in this study. It is unlikely that malaria episodes recurring from day15 onwards were recrudescences as the standard doses of SP given would elicit drug concentrations in vivo that are in excess of those required to inhibit sensitive P. falciparum strains found in East Africa (15,16). This implies that unless reinfection is prevented throught the RSP span (e.g by insecticides treated nets), selection for resistant strains is unavoidable once SP is in widespread use. The prevalence of DHPS mutations was very low compared to DHFR mutations, and the pre-treatment DHPS mutation at codon 436 was the most frequent, a common observation in areas with least in vivo SP resistance (17). The DHPS mutation at codon c437 occurred more frequently in parasites sampled during the RSP span than before treatment indicating possible selection for this mutation, which is common in areas with widespread SP use and resistance (17). Studies on the prevalence of DHFR & DHPS mutations in areas with widespread SP use and resistance show that mutations in both enzymes are associated with SP resistance (18). The study has shown that, despite supervised SP administration with optimal dosage, in areas with high intensity of malaria transmission, there is a very high probability of in vivo selection for parasites with mutations at re-infection because of the long terminal elimination half-life of SP (1,14). In routine use as a 1st line drug, no matter how well prescribed and the degree of compliance, selection for resistance is unavoidable, and since SP induces gametocytaemia (19), parasites carrying mutations will be transmitted and maintained in the natural population of P.falciparum infections, and be subjected to continuous SP selection in which case resistance would build up rapidly (20) as is the case now in most of the Eastern and South African countries for which artemisnin based combination therapy is the option (21). Acknowledgement This work was supported by DANIDA / ENRECA under the Tanzanian – Danish Collaborative Research and Training Programme, Phase III 1998 – 2001, Project III and partly by SAREC under the Muhimbili – Karolinska Collaboration. Thanks go to the Permanent Secretary, Ministry of Health Tanzania for approving the study. Special thanks go to Mr Frederick Kalokola, for tirelessly handling the laboratory work in Tanzania and Mr Jimmy Weng for technical assistance on the DHFR/DHPS genotyping in Copenhagen. Thanks are due to the mothers and staff of Kibaha Hospital for their co-operation in this study. This work is dedicated to the late Jeremiah Masunga (C.O) for working with me tirelessly. References
© Copyright 2005 - East African Journal of Public Heath The following images related to this document are available:Photo images[lp05015t2.jpg] [lp05015t3.jpg] [lp05015t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}