 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Indian Journal of Medical Microbiology, Vol. 23, No. 1, January-March, 2005, pp. 14-19 Special Article Human anti-rhinosporidial antibody does not cause metabolic inactivation or morphological damage in endospores of Rhinosporidium seeberi, in vitro Arseculeratne SN, Atapattu DN, Eriyagama NB Department of Microbiology, Faculty of Medicine, University
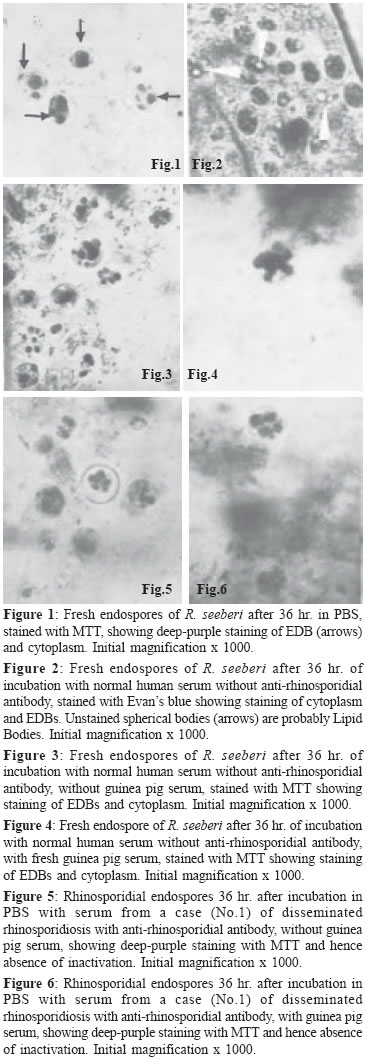
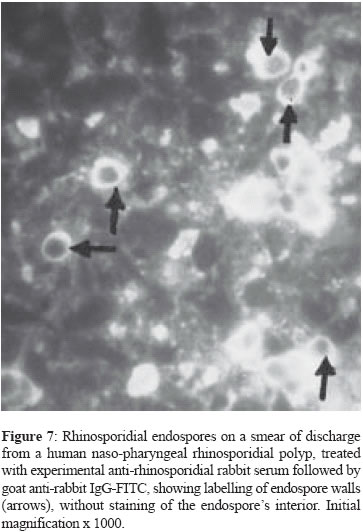
of Peradeniya, Peradeniya Code Number: mb05003 ABSTRACT This report describes the use of the MTT-reduction and Evan's blue-staining tests for the assessment of the viability and morphological integrity, respectively, of rhinosporidial endospores after exposure to sera from rhinosporidial patients with high titres of anti-rhinosporidial antibody. Sera from three patients, with nasal, ocular and disseminated rhinosporidiosis respectively were used, with human serum without anti-rhinosporidial antibody for comparison, with or without added fresh guinea pig serum as a source of complement. All four sera tested, with or without guinea-pig serum, had no effect on the morphological integrity or the viability of the endospores and it is suggested that anti-rhinosporidial antibody has no direct protective role against the endospores, the infective stage, in rhinosporidiosis. This finding is compatible with the occurrence of chronicity, recurrence and dissemination that are characteristic of rhinosporidiosis despite the presence of high titres of anti-rhinosporidial antibody in patients with these clinical characteristics. The possible occurrence of humoral mechanisms of immunity that involve anti-rhinosporidial antibody with cells such as leucocytes and NK cells, in vivo, cannot yet be discounted, although the presence of high titres of anti-rhinosporidial antibody in patients with chronic, recurrent and disseminated lesions might indicate that such antibody is non-protective in vivo.Key words – Rhinosporidium seeberi, anti-rhinosporidial antibody Rhinosporidiosis in humans presents several enigmas that have yet to be explained: (a) the chronicity of the disease that could last for several years, (b) recurrence after surgical excision of the initial polyps that most commonly occur in the anterior respiratory passages, and (c) occasional dissemination from these sites to the limbs and viscera. Cell-mediated immune (CMI) responses were recently demonstrated by the assay of lymphoproliferative responses to rhinosporidial antigens and the T-cell mitogen Concanavalin A of peripheral blood lymphocytes of patients with rhinosporidiosis[1] and by the foot-pad delayed-type hypersensitivity response to rhinosporidial antigens in experimental mice.[2] The occurrence of CMI responses was also suggested by the marked presence in rhinosporidial polyps of CD8+ lymphocytes and TIA1-containing CD8+ and CD57+ NK cells, with numerous macrophages; these host-cell infiltrates were often associated with degenerate rhinosporidial sporangia.[1] The role of antibody mediated anti-rhinosporidial immunity has however not been investigated although high titres of anti-rhinosporidial antibody have been detected in patients with localized or disseminated disease.[3] The lack of a method for in vitro culture of Rhinosporidium seeberi and for the establishment of a model of rhinosporidiosis in experimental animals, has contributed to the paucity of studies on the role of immune mechanisms in recovery from rhinosporidiosis. There are also, hitherto, no standardised methods described for the assessment of the viability of rhinosporidial ontogenic elements on which studies on the protective effect of CMI and anti-rhinosporidial antibody could be made in vitro. There has been only one report on the viability assessment of rhinosporidial endospores that stated that the Nigrosin-Eosin stain was capable of differentiating between living and dead endospores.[4] The monotetrazolium salt MTT (3-4],5-dimethylthaizol-2-yl]-2,5-diphenyltetrazolium bromide) has been used to assess the viability of mammalian cells, especially lymphocytes[5],[6],[7] and of mycelial and yeast fungi.[8],[9] This method depends on the production of a deep purple-coloured formazan by the reduction of the MTT by mitochondrial dehydrogenases of viable cells. This method was used by Levitz and Diamond[8] to assess the sensitivity of mycelial fungi to hydrogen peroxide, and by Jahn et al[9] to assess the susceptibility of Candida albicans and Aspergillus spp. to anti-fungal agents. We reported[10] that MTT could differentiate between inactivated, dead endospores of Rhinosporidium seeberi, after heating (100°C or autoclaving), or treatment with 10% formalin from viable endospores with the finding that only the latter produced the formazan derivative in both the endospore′s cytoplasm and in a proportion of the spherical bodies within the endospore that were provisionally identified as the electron dense bodies - EDBs.[11] We have also used the MTT method to assess the sensitivity of rhinosporidial endospores to biocides [12] and to anti-microbial drugs.[13] This report describes the application of the MTT and Evan′s blue-staining methods to the investigation of the effects of human sera from rhinosporidial patients, with or without added guinea pig serum as a source of complement, on the metabolic activity and morphological integrity of rhinosporidial endospores in vitro; untreated endospores and those treated with anti-rhinosporidial serum, with or without added complement, showed normal staining with MTT and with Evan′s blue. We conclude that neither sera from normal (non-rhinosporidial) persons nor anti-rhinosporidial sera from patients with rhinosporidiosis that contained anti-rhinosporidial antibody inhibited MTT-reduction by the endospores and hence anti-rhinosporidial antibody was incapable of inactivating the endospores; this result is in accordance with the occurrence of the predominant clinical characteristics of rhinosporidiosis - chronicity, recurrence and dissemination despite the presence of high titres of anti-rhinosporidial antibody.[3] MATERIALS AND METHODS Buffers 0.2M phosphate buffered-saline (PBS) at pH 7.2 was used to suspend endospores, to dilute the normal and rhinosporidial sera, and to dissolve MTT and Evan′s blue. Endospores Only endospores were examined as the possible target of antibody-mediated destruction because (1) these are considered to be the infective stage of R. seeberi that gains access to susceptible sites from the pathogen′s natural habitat - ground water, and because "auto-inoculation"[14] occurs on mucosal surfaces by endospores when they are released from traumatised or excised polyps leading to the formation of satellitic rhinosporidial lesions, (2) sporangia have a pore for the exit of mature endospores, through which passive ingress of the dyes might have occurred; (3) MTT-staining of sporangia was observed only in the endospores they contain. Fresh endospores were obtained by homogenising nasal rhinosporidial tissues, soon after surgery, in distilled water to lyse osmotically labile host cells. The homogenate was centrifuged to deposit the endospores, the deposit was washed once in distilled water and the final pellet re-suspended in PBS to a density of approximately 2.5 x 106 endospores/mL as counted in an improved Neubauer blood cell counting chamber; a few developing endospores that were in the stage of early juvenile sporangia were present. Immature and mature sporangia were absent; they were probably ruptured by the homogenisation. The mature sporangia would have released their content of endospores on rupture. Test sera Sera from three human patients with rhinosporidiosis, stored at - 20°C, were used in comparison with serum from a 1 year-old infant (obtained from a serum bank) that showed no anti-rhinosporidial antibody. The rhinosporidial sera showed the following titres on dot-blot ELISA assays with anti-human immunoglobulin-phosphatase conjugates[3] - Disseminated rhinosporidiosis- IgG 1/640, IgM 1/60 Incubation with sera The normal and rhinosporidial sera were diluted 1/10 in PBS and used without heat-inactivation or added fresh, guinea pig serum as a source of complement. In other experiments, the unheated sera were used at the same dilutions in PBS containing fresh 1/10 normal guinea pig serum. To the centrifuged deposit from approximately 50 µL of the endospore suspension in 1.5 mL, capped, plastic microfuge tubes, was added approximately 150 µL of the diluted serum, followed by side-tapping of the tube for dispersion of the endospores. The tubes were incubated at room temperature (28°C ambient) for 36 hours. Endospores, deposited from the original suspension, re-suspended in PBS and incubated in a similar manner were used as controls. The MTT test In the assessment of lymphocyte proliferation, MTT is used at a concentration of 5 mg/mL, which is diluted to 0.5 mg/mL when the dye-solution is added to the wells containing growth medium and lymphocytes. Since, in our tests, the dye-solution was added to the supernatant-free deposit of endospores, the concentration of MTT was reduced to 0.5 mg/mL. The MTT solutions were freshly prepared. After incubation, the tubes were centifuged at 400 x g at room temperature (RT), and the endospore-pellets were washed thrice with PBS. To the pellets were added approximately 50 µL of MTT (0.5 mg/mL), the mixtures were agitated by side-tapping of the tubes and then incubated at 37°C in air for 3 hr. After incubation, a drop of the suspension was placed on a microscope slide under a cover-slip for microscopy at x 1000, under oil. The intensity of the deep purple formazan-staining in the cytoplasm and the spherical bodies (EDBs) within the endospore and the distribution of positively-stained endospores were assessed qualitatively and taken as indices of metabolic activity of the endospores. Quantitative counts of staining were not done as the host tissue cells in the endospore-suspension obscured some endospores. In addition to the intensity of staining, the morphology of the endospores (walls and spherical bodies) were also observed. The Evan′s blue test Two drops of 0.2 % Evan′s blue in PBS were added to the washed pellets of control (untreated) or serum-treated endospores, and the mixtures after agitation, were incubated at RT for 1 hr. A drop of the mixture was then placed on a microscope slide under a cover-slip for examination at x 1000 under oil. RESULTS Control, serum-free endospores showed intense purple-staining of their EDBs with MTT, indicating their metabolic activity and hence viability after 36 hour of incubation in PBS [Figure - 1] in over 95% of the endospores. With Evan′s blue the cytoplasm showed moderate staining while the EDBs stained deeply in endospores incubated with normal serum [Figure - 2]. Endospores incubated in normal human serum without added guinea pig serum [Figure - 3] and endospores incubated in normal human serum with added guinea pig serum [Figure - 4] showed deep staining of the EDBs and the cytoplasm with MTT in a similar proportion as in untreated endospores. Endospores that were exposed to a human serum that contained high titres of anti-rhinosporidial antibody without added guinea pig serum [Figure - 5] or with added guinea pig serum [Figure - 6] showed deep staining of the EDBs and cytoplasm. The intensities of the staining of the EDBs in the control endospores and its distribution, as well as of the serum-treated endospores were similar indicating that neither the anti-rhinosporidial antibody-free human serum nor the high-titred anti-rhinosporidial serum inactivated the endospores. Negative results were obtained with all 3 sera from patients with rhinosporidiosis. The intensity of staining with Evan′s Blue of the cytoplasm and the EDBs of the endospores exposed to normal (anti-rhinosporidial antibody-free) serum or to sera that contained anti-rhinosporidial antibody showed no differences from that of the control endospores, indicating, as with the MTT stain, that no morphological abnormalities were produced by any of the sera tested. DISCUSSION In the absence of a method for the in vitro culture of R. seeberi, its "life-cycle" has been deduced from the appearances of the different ontogenic stages of the pathogen in the rhinosporidial granulomata of diseased hosts. The endospore that contains a thick wall with chitin and cellulose, develops into the thin-walled juvenile sporangium (trophocyte) from which arise the immature (intermediate and mature) sporangia. Endospore walls are different in structure and function from the walls of the cysts of parasites and the endospores of bacteria that are resting, non-vegetative stages. The rhinosporidial endospores thus retain semi-permeability to nutrients and small molecules of dyes such as MTT (MW 414). Endosporulation in the mature rhinosporidial sporangia leads to the formation of thousands of endospores which repeat the life cycle when released from the mature sporangium. None of the conventional in vitro culture methods has succeeded in providing serial propagation of the ontogenic stages and hence no culture methods are available for assessment of their viability; hence we used the indirect method of MTT-staining for this purpose. Evan′s blue was earlier shown[10] to stain the cytoplasm and, more intensely, a proportion of the spherical bodies within the endospore, that were regarded as the electron dense bodies - EDB.[15],[16] Spherical bodies that did not stain were regarded as the lipid bodies (LB). MTT was shown to stain areas in the cytoplasm as well as some of the spherical bodies, that were regarded as the EDB while the MTT-negative bodies were probably the LB.[11] The reduction of MTT to the purple-coloured formazan depends on mitochondrial dehydrogenases of living cells and hence the formazan production within the endospores was interpreted as indicating their viability. Levitz and Diamond[8] used MTT to assess the effect of hydrogen peroxide on mycelial fungi and demonstrated that MTT-negativity correlated with failure of growth of the peroxide-treated fungi in culture. Jahn et al[9] used the MTT method to demonstrate that the effects of antifungal agents on Candida albicans correlated well with MIC determinations. The retention of deep MTT-staining by the contents of the endospores despite treatment with anti-rhinosporidial antibody, with or without added complement in our experiments, is therefore interpreted by us to mean that such antibody has no inactivating, devitalising effect on rhinosporidial endospores; the morphological integrity of the endospores and their EDBs as also shown by staining with Evan′s blue, further indicates that anti-rhinosporidial antibody does not cause morphological damage to the endospores. Some parallels probably exist between R. seeberi and Toxoplasma. The encysted bradyzoites of Toxoplasma persist for the life-time of the host,[17] presumably evading destruction by anti-Toxoplasma antibody that can however kill the extracellular, non-encysted form.[18] The cyst that contains the bradyzoites is "surrounded by a dense argyrophilic wall enclosed within the host-cell cytoplasm"[19] and is thereby probably inaccessible to antibody; the bradyzoite cyst wall probably acts in a manner similar to that of the rhinosporidial endospore′s wall in excluding the penetration of antibody. A further parallel of toxoplasmosis with rhinosporidiosis is that despite high circulating antibody titres, immunosuppressed patients develop disseminated toxoplasmosis[19] recalling the fact that despite high anti-rhinosporidial antibody titres, dissemination occurs in rhinosporidiosis; in toxoplasmosis and rhinosporidiosis, the corresponding antibodies probably do not penetrate into the cyst and the endospore respectively. A further explanation of the failure of anti-rhinosporidial antibody to inactivate the endospores of R. seeberi, exists in a previous report that described the presence of human immunoglobulin bound to soluble antigens released by sonication of rhinosporidial endospores and sporangia, and detected by immuno-blots and SDS-PAGE.[20] It was pointed out that such binding of host-protein could be a mechanism of evasion of immune attack by anti-rhinosporidial antibody in diseased hosts.[21] This binding-phenomenon might also explain the absence of an inactivating effect of human anti-rhinosporidial sera on the endospores, as found in the present study. A similar phenomenon has been described of schistosomes in which older parasites have bound host-proteins; "They are only susceptible to attack by specific antibody in vitro if they are not coated with host protective antigens".[22] On the other hand, while binding was not found with rabbit immunoglobulin, endospores in smears from rhinosporidial polyps were found[23] to be labelled with experimental anti-rhinosporidial rabbit serum traced with FITC-labelled anti-rabbit goat serum [Figure - 7] indicating that endospores are antigenic and that they are reactive on their surface with anti-rhinosporidial serum that contained anti-endospore antibody. However it is noteworthy that [Figure - 7] also illustrates the staining of the endospore wall without staining the interior of the endospore; this failure might be attributable to the non-penetration of the antibodies and the anti-rhinosporidial serum and tracer antibody-conjugate into the endospore′s interior, as perhaps with the endospores that failed to be inactivated in the tests with anti-rhinosporidial antibody. Our in vitro results suggest that anti-rhinosporidial antibody had no effect (sporicidal or sporolytic) on the viability and integrity of rhinosporidial endospores; this finding might imply the failure of such antibody to directly inactivate the endospores in vivo in infected tissues. On the other hand other anti-rhinosporidial antibody-dependent mechanisms[24] such as immune opsonization, intra-phagocytic and extra-phagocytic killing, and antibody dependent cell-mediated cytotoxicity could conceivably operate in vivo; the neutralization by antibody of soluble rhinosporidial products might also aid the process of resolution. Yet the presence of high titres of anti-rhinosporidial antibody in patients with chronic, recurrent and disseminated rhinosporidiosis[3] indicates that anti-rhinosporidial antibody is not protective. The apparent ineffectiveness of anti-rhinosporidial antibody-containing sera from patients in inactivating endospores in vitro, might need further examination in view of recent findings on anti-mycobacterial antibody. In a reconsideration of the role of anti-Mycobacterium tuberculosis antibody in protection, Glatman, Freedman and Casadevall[25] pointed out that polyclonal serum preparations have had no consistently-demonstrable protective activity against some fungal infections, caused by Cryptococcus neoformans, Candida albicans and Pneumocystis carinii, but that monoclonal antibodies do have protective activity, with the possibility that in natural infections, a mixture of polyclonal antibodies with protective, non-protective or disease enhancing activities exists. It is a conceivable parallel that in rhinosporidiosis a similar effect operates, accounting for both the failure of antibodies in diseased patients to provide protection and polyclonal anti-rhinosporidial sera in these patients to produce inhibition of R. seeberi, in vitro. ACKNOWLEDGEMENT We thank Dr P Balasooriya and Dr S Senanayake for supplies of excised rhinosporidial polyps, Dr NR de Silva for a gift of MTT. REFERENCES
Copyright 2005 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb05003f7.jpg] [mb05003f1-6.jpg] |
| |||||||||
{kind=link}
{kind=link}