 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
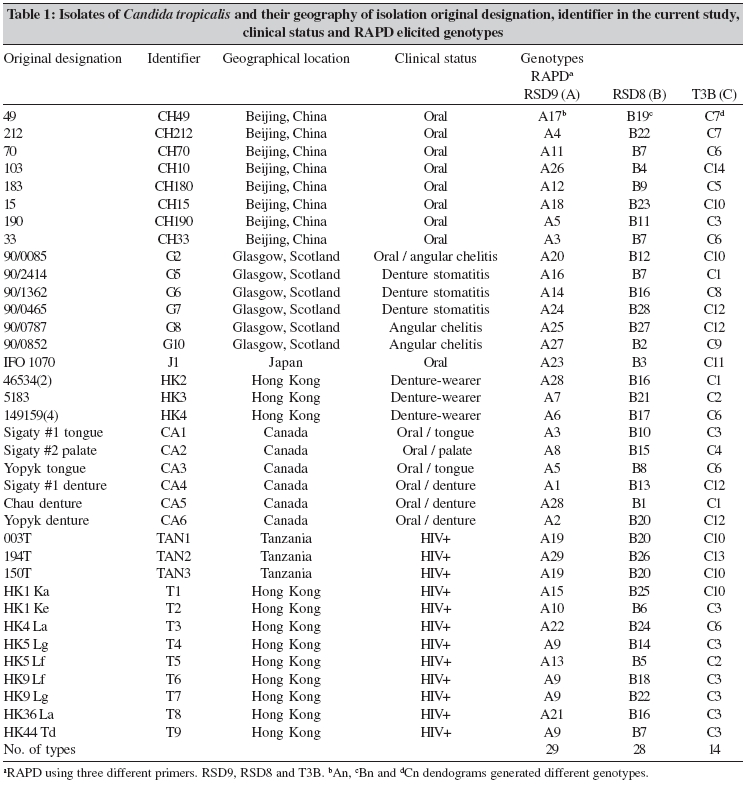
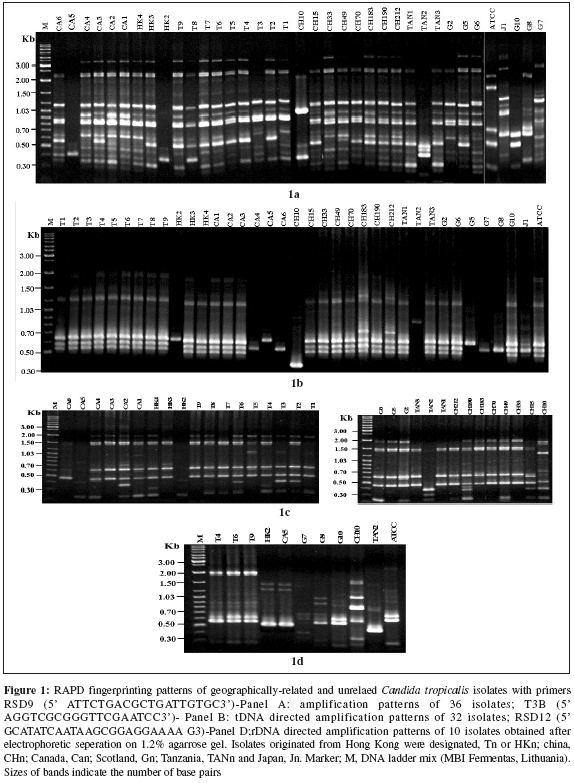
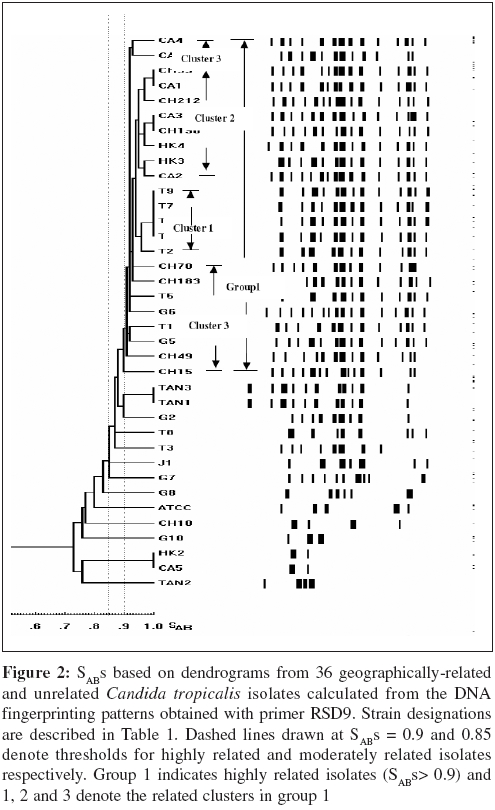
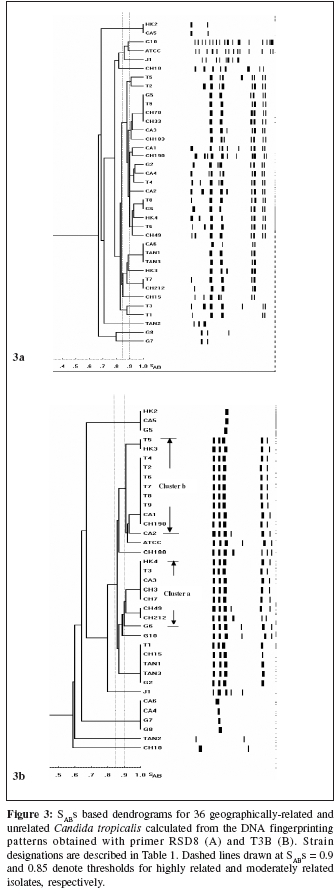
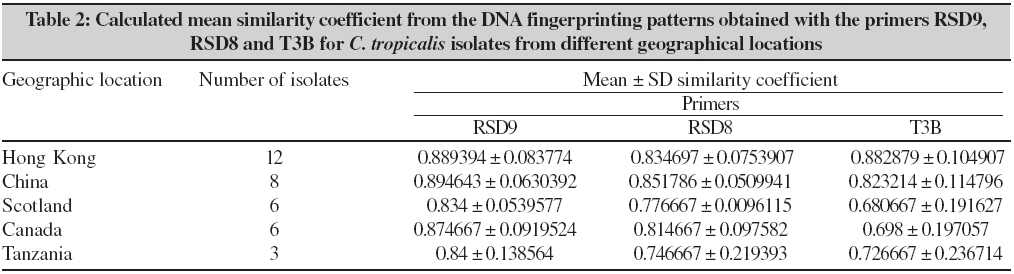
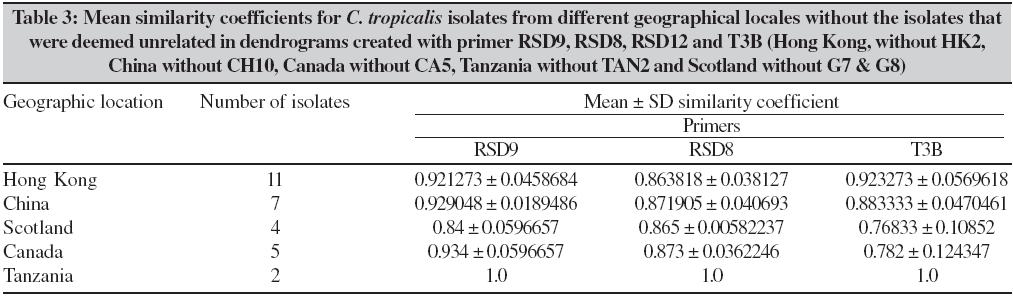
Indian Journal of Medical Microbiology, Vol. 24, No. 3, July-September, 2006, pp. 186-194 Original Article DNA fingerprinting elicited evolutionary trend of oral Candida tropicalis isolates from diverse geographic locales Dassanayake RS, Samaranayake YH, Yau JJY, Samaranayake LP Faculty of Dentistry, University of Hong Kong, Hong Kong SAR Code Number: mb06056 Abstract Purpose: To study molecular profiles of oral Candida tropicalis isolates from five different geographic locales to determine the molecular diversity, clonality and evolutionary trends of this opportunistic pathogen.Methods: A total of 36 strains from five countries (China, Canada, Scotland, Japan and Tanzania) were genotyped by PCR fingerprinting with 11 separate primers. Of these, primers RSG9, RSG8, T3B and RSD12 generated complex fingerprinting patterns. Results: Three significantly dissimilar profiles were derived from the primer T3B and particularly focused on tDNA suggested the prevalence of genetic subtypes within the species. Comparison of tDNA and rDNA (RSD12) fingerprints of C. tropicalis suggested that rDNA is much more heterogeneous than the relatively distinct tDNA. Further analysis of similarity coefficient (SAB) of gel profiles derived from computer-generated dendrograms indicated some degree of similarity in isolates from five-disparate geographic locales as well as the presence of unique isotypes in each region. Conclusions: This study demonstrates the evolutionary divergence of distinct genetic subgroups within Candida tropicalis . Keywords: Candida tropicalis, evolution, fingerprinting, PCR Opportunistic pathogen Candida causes common human fungal infections that manifest both superficially and systemically, especially in compromised patients. Although, C. albicans is by far the main aetiological agent of candidosis, the frequency of isolation of other non- albicans species is increasing at an alarming rate. Candida tropicalis is the second most common Candida species colonizing humans[1],[2] and a major cause of invasive candidal infection.[3],[4] This organism has become a significant health problem as a nosocomial agent especially in patients with haematological malignancies and cancer[5] or those undergoing bone marrow transplantation.[3],[6],[7] It also causes systemic disease[8] and chronic volvovaginitis.[1] In a recent study, the death of a patient due to C. tropicalis endocarditis has been reported, where herniated fungal vegetations completely obstructed the flow of the ascending aorta.[9] Interestingly the yeast is often resistant to fluconazole, a popular triazole agent that is used globally.[10] To date, the molecular epidemiology of C. tropicalis using DNA-based typing methods has been sparsely studied. Lin and Lehmann[11] employed a PCR based fingerprinting method to show that nine epidemiologically related isolates from an outbreak of sternal wound infection were either identical or very similar. Recently, Correia et al[2] employed latter method to study molecular epidemiology of clinical isolates of C. tropicalis and other Candida species derived from two medical institutions in Portugal and showed that the number and size of the amplification products were characteristic for each species. Vrioni and Matsiota-Bernard[12] also adopted similar approach to demonstrate that urinary tract colonization/ infection by C. tropicalis is both endogenous and exogenous in an intensive care unit in the US. In another study, a total of 89 clinical isolates of C. tropicalis from 56 patients from four different medical centers in U.S. were analyzed using pulse-field gel electrophoresis following restriction digestion of genomic DNA yielding 49 different DNA types.[4] In addition, restriction fragment length polymorphism[13] and Southern blot analysis of restriction digested genomic DNA with a moderately repetitive, species-specific DNA probe[7] have also been used to type C. tropicalis . The foregoing investigators have indicated that molecular fingerprinting methods that effectively investigate subtypes of C. tropicalis are vital for understanding the clinical epidemiology, emergence of resistance strains during antifungal therapy, nosocomial outbreaks of infection and evolutionary and taxonomic diversity. However, to our knowledge detailed investigations of C. tropicalis subtypes especially in relation to their geographic origin has not been undertaken. In our previous studies we have found that the RAPD technique is a promising tool for yeast genotyping, especially when used with different primer combinations[14],[15],[16] and the results are amenable to computer assisted cluster analysis.[16] Therefore, the aim of the current study was to investigate the RAPD fingerprints of 36 C. tropicalis isolates from varying geographical locales using tDNA and rDNA directed sequence-specific and arbitrary primers in order to asses their genomic heterogeneity. Further, an attempt was made to investigate their intraspecies genetic relatedness as a recent report has hinted the possible existence of a genetic variant of yeast which may be classified as a new species.[17] Materials and Methods C. tropicalis isolates A total of 36 oral C. tropicalis from different geographic locales was used in this study. They originated from Hong Kong (designated as Tn or HKn), China (CHn), Canada (CAn), Scotland (Gn), Tanzania (TANn) and Japan (Jn). The yeasts were derived from human immunodeficient virus infected patients, denture-wearers, patients with angular chelitis, denture stomatitis, burning mouth syndrome and black hairy tongue [Table - 1]. Candida tropicalis ATCC 13803 was used as a control. The organisms were identified using the commercially available API 20C Candida identification kits (API system, Vercieu, France). After recovery the yeasts were maintained on Sabouraud dextrose agar (SDA) and stored at 4-6°C during the experimental period. Genotypic characterization Preparation of DNA for randomly amplified polymorphic DNA (RAPD) analysis. Yeast obtained from stock cultures were subcultured on yeast-peptone dextrose (YPD) medium (1% peptone, 1% yeast extract, 2% glucose, 1.5% agar) at 37°C for 24 hours and single colonies were transferred to 20 mL of YPD broth (1% peptone, 1% yeast extract, 2% glucose) and incubated at 30°C under aerobic conditions to the stationary phase (as assessed by the measurement of the optical density of the culture at 600 nm). Following incubation, yeasts were harvested by centrifugation at 4000 rpm for five minutes and washed in 1M Sorbitol. The yeast pellet was resuspended in 1.5 ml SE buffer (1.2 M sorbitol, 0.1 M EDTA pH 8.00) containing 3 mL of β -mercaptoethanol (Sigma Chemical Co., Louise, MO) and 0.5 mg yeast lytic enzyme (Lyticase, Sigma Chemical Co., Louise, MO) incubated at 37°C for at least one hour until formation of spheroplasts and harvested by centrifugation at 13,000 rpm for five minutes. These spheroplasts were washed twice in SE buffer and resuspended in 1.5 mL of 0.15M NaCl, 0.1M EDTA pH 8.00 and lysed by addition of proteinase K (final concentration 500 µg/ mL) and, SDS (1% (W/V) final concentration) followed by the addition of RNase (500 µg /mL) and incubated at 55°C for one hour. The resulting supernatant obtained following centrifugation at 13000 rpm was extracted twice with phenol and once with phenol/chloroform prior to precipitation of DNA by addition of equal volume of 2-propanol. The DNA precipitated was finally dissolved in 100 μL of TE buffer (10 mM Tris, 0.1 mMEDTA pH 8.0). Randomly amplified polymorphic DNA (RAPD) analysis The custom synthesized primers (Gibco BRL, Hong Kong) used in this study were OBU1: 5′CACATGCTT3′, OBU2: 5′CACATGCCT3′, OBU3: 5′CGCATGCTT3′, RSD6: GCGATCCCCA3′, RSD8: 5′GGTCCGTGTTTCAAGACG3′, RSD12: 5′GCATATCAATA AGCGGAGGAAAAG3"; RSD7: 5′AGTGAATTCGCGGTGAGATGCCA3′; RSD11: 5′A ACGCGCAAC3′; RSD9: 5′ATTCTGACGCTGATTGTGC3′; RSD10: 5′CCGCAGC CA3′ and T3B: 5′ AGGTCGCGGGTT CGAATCC3′.[14],[15],[16] Thermocyling was performed in a GeneAMP 9700 machine (Perkin Elmer, Foster City, USA). PCR master mix containing approximately 20 ng of yeast DNA as template, 5 μ L of 10x PCR buffer (200 mM Tris-HCl (pH 8.4) and 500 mM KCl), 200 mM of dNTPS, 2 mM MgCl2, 1.5 mM of primer, 1.5 U Taq polymarase (Life Technologies, USA) in 50 mL was used for PCR reaction. The first 5 cycles included 30 seconds of denaturation at 94°C, 2 minutes of annealing at 21°C (primers OBU1, OBU2 and OBU3), 23°C (RSD6 and RSD10) 45°C (primers RSD11 and RSD12), 43°C (RSD9 and RSD8), 47°C (RSD7) and 41°C (T3B) and 2 minutes of primer extension, followed by 45 cycles of 30 seconds of denaturation at 94°C, 2 minutes of annealing at 27°C (primer OBU1, OBU2, OBU3), 29°C (RSD6 and RSD10), 52°C (RSD11 and RSD12), 49°C (RSD9 and RSD8), 54°C (RSD7) and 47°C (T3B), 2 minutes of primer extension at 72°C. The reaction was held at 72°C for 15 minutes. Control tubes containing master mix but without template DNA were included in each run and reproducibilty checked for each reaction. The PCR products were electrophoresed in agarose gel (1.2%) for approximately 2 hours at room temperature in 0.5 TBE buffer (89 mM Tris, 89 mM-boric acid, 2.5 mM EDTA, pH 8), stained with ethidium bromide and visualized with UV light.[15] Analysis of fingerprints RAPD gel profiles captured by a digital camera were then digitized using the Dendron software package version 2.0 (Solltech, Iowa City, Iowa). "Unwrapping option" of Dendron was used to straighten distorted gels and "detect lanes and bands options" used to automatically identify the lanes and bands. The similarity coefficient (SAB) computation based on band position alone for every pair of isolate profiles was calculated using an algorithm. In generating the algorithm the data for two banding patterns (lane A and B) were synopsized by the binary value 0 and 1, where 0 indicates no band at position and 1 indicates a band at that position. SAB for each pair of strains, A and B, was calculated by the formula SAB = 1- √ b+C/2a+b+c. Where a is the number of bands common for both lane A and B (coded as 1, 1), b is the number of bands in lane A with no counterpart in lane B (coded as 1, 0), c is the number of bands in lane B with no counterpart in lane A (coded as 0,1). Dendrograms based on SAB were automatically created with the Dendron software programme by the unweighted pair group method.[18] The gels were normalized in order to avoid inter and intragel variation and aligned by associating bands of internal molecular weight standards on each gel with stored reference positions. Isolates with identical RAPD profiles were given a SAB value of 1.00, whereas SAB of 0.00 was given for non-identical isolates having RAPD profiles with no correlating bands. Thus SAB values ranging from 0.01 to 0.99 denoted profiles with increasing number of congruent bands between RAPD profiles. On interpretation of results of SAB values 0.71, 0.85 and 0.90 were used as cut-off points to segregate cluster of unrelated, moderately related and related strains undergoing microevolution, respectively.[19] Results Generation of discriminatory fingerprints using nonspecific and sequence-specific primers In initial PCR studies, genomic analyses of 36 C. tropicalis isolates were performed using 11 different primers Table - 1. However, only the fingerprints generated by primers RSD8, RSD9, RSD12 and T3B were adequately discriminatory [Figure - 1] for the current purpose and hence used in the remainder of the study. The PCR protocols used for different primers were optimized with regards to the annealing temperature whilst other parameters were kept constant. The primer RSD9 produced upto 22 polymorphic bands and exhibited the highest discriminatory power defining 29 genotypes (A1-A29; [Table - 1]; [Figure - 1]a and[Figure - 2]) with a mean similarity coefficient (SAB ) value of 0.86, whereas primers RSD8 produced up to 20 polymorphic bands generating 28 genotypes (B1-B28; [Table - 1]; [Figure - 1]c and [Figure - 3]a) with a mean SAB , value of 0.81. The tDNA directed primer T3B generated nearly 12 polymorphic bands thus sub dividing 36 C. tropicalis isolates into 14 genotypes (C1-C14; [Table - 1]; [Figure - 1]b and [Figure - 3]b) with a mean SAB , value of 0.79. Strains from same geographical locale are related but not always genetically distinct Although some isolates from different geographical locales did generate divergent genomic profiles, the majority of isolates from Hong Kong, China, Canada, Scotland and Tanzania were related [Table - 2]. This was evident when separate dendrograms were created with the PCR fingerprinting patterns generated from primers T3B, RSD8 and RSD9. Apart from a single exception, an isolate from Hong Kong (HK2), all other isolates generated a genetically similar cluster [Table - 3]. A similar trend was observed in isolates from other geographical locales yielding genetically similar isolates with a few exceptions. However, it was evident the isolates that originated from Scotland were significantly different with mean similarity coefficients that were much lower than those from other locales [Table - 2][Table - 3]. This genomic disparity was quite conspicuous when these isolates were fingerprinted with primer T3B [Figure - 3]b, [Table - 1]. On examination of the dendrograms related to the primer RSD9, we identified a group of highly related isolates (group 1, [Figure - 2]), which can be further divided into three major clusters on scrutiny. The cluster 1 comprised either identical or highly related isolates (T9, T7, T6, T4 and T2) with SABs ~0.95 or greater; cluster 2 consisted of identical and highly similar strains (CH33, CA1, CH212, CA3, CH190, HK4, HK3 and CA2) with a mean SAB s of 0.95. However, the cluster 3 represented similar but non identical isolates (CA4, CA6, CH70, CH183, T5, G6, T1, G5, CH49 and CH15) with SABs of 0.9. When combined dendrograms were generated separately using the fingerprinting patterns of primers T3B, RSD12 and RSD8, all the isolates in cluster 1 from Hong Kong, were found to be identical (SAB s 1) with primer T3B [Figure - 3]b. Further with primer RSD12, the isolates in the latter cluster behaved similarly either remaining identical (T6, T7 and T9) or highly related (T2 and T4) with SABs of 0.94 or above [Figure - 4]. However, with the primer RSD8 the isolates from this cluster were randomly distributed with others from different geographic regions [Figure - 3]a. Thus, it appears that the isolates from Hong Kong form a genetically distinct cluster at least with three of the primers used. On investigating the genomic profiles of cluster 2 isolates (CH33, CA1, CH212, CA3, CH190, HK4, HK3 and CA2) [Figure - 2] with primer RSD8, CH33, CA1, CA3, CH190, HK4 and CA2 isolates remained in a single cluster with SABs of 0.9 or greater with random mixing [Figure - 3]A. However, with primer T3B these isolates separated into two different clusters with SAB s of 0.9 or above (cluster a: CH33, CH212, CA3 and HK4, cluster b: CA1, CH190, HK3 and CA2) [Figure - 3]b. This analysis revealed that the isolates CH33, CA3 and HK4 and CA1, CH190 and CA2 are genetically closely related. On examining the dendrogram of the fingerprints of primer RSD8, some of the isolates (T5, G5, CH70, CH183, CA4, G6 and CH49) of cluster 3 remained in a cluster with SAB s of 0.9 or above with other penetrating strains [Figure - 3]a. However, with primer T3B, cluster 3 isolates split into two clusters with SAB s of 0.9 or greater with other penetrating strains. The isolates CH49, CH70 and G6 remained in one cluster whereas isolate T5 fall into second cluster [Figure - 3]b. This was the case with primer RSD12, where isolates CH49, CH70 and G6 fell into a cluster and T5 stayed in a related cluster [Figure - 4]. Overall analysis of cluster 3 isolates indicated that isolates CH70, G6 and CH 49 are highly related and this group is genetically similar to isolate T5. These analyses revealed that some isolates from diverse geographic locales (Hong Kong, Canada, China and Scotland) are genetically homogeneous. Nonetheless, further investigation of dendrograms generated with primers T3B, RSD9 and RSD8 indicated that yeast isolates from the same geographic locale were also similar. For instance in the case of the isolates TAN1 and TAN2, which originated from Tanzania, remained identical [Figure - 1][Figure - 2][Figure - 3]a and b. The presence of genetically divergent isolates within the species of C. tropicalis As demonstrated above, the PCR fingerprinting of C. tropicalis with primer RSD9 and the genesis of dendrogram revealed that the isolates in cluster 1, 2 and 3 are genetically similar (mean SAB - 0.9, group 1), while a group of other isolates (CH10, CA5, HK2, TAN2, G8 and G7), which produced distinct fingerprints remained unrelated with SABs of 0.84 or lower [Figure - 2]. A similar trend was observed in PCR fingerprints of the latter group with other three primers. Thus they remained distinct with the SABs of ≤0.65, ≤0.80 and ≤0.71 or lower with primers T3B, RSD8 and RSD 12, respectively [Figure - 1][Figure - 3][Figure - 4]. tDNA directed PCR fingerprinting profiles divided isolates into three major subgroups As the number and sizes of tDNA directed PCR fingerprinting patterns are characteristic for a particular species, we utilized this approach to identify the existence of subspecies or hidden species of C. tropicalis . The tDNA PCR fingerprinting revealed three major subprofiles of C. tropicalis . Multiple bands predominate in one of these whilst the other two profiles showed a single intense band of differing size [Figure - 1]b and [Figure - 3]b. Most of the latter isolates were those that produced divergent RAPD profiles with other primers. Isolates from HIV infected individuals are similar to those from the healthy individuals An attempt was made to study the relationship between C. tropicalis isolates from HIVinfected and healthy individuals by comparing the dendrograms generated from fingerprinting patterns with primers T3B, RSD8 and RSD9. In this manner isolates from HIV-infected individuals of Hong Kong and Tanzania were compared with those from healthy individuals in Hong Kong, China, Canada and Scotland; the calculated mean SAB s values revealed no significant difference between the healthy and HIV-infected groups [Table - 4]. Discussion C. tropicalis is a major agent of invasive candidal infection and the third most frequent Candida species colonizing the oral cavity after C. albicans and C. glabrata. Some strains of C. tropicalis are resistant to fluconazole, a widely used highly efficacious antimycotic agent.[10],[11] Despite these reports emphasizing the increasing clinical significance of C. tropicalis , there are only a few DNA-based studies that have attempted to characterize this ubiquitous yeast and, no studies as far as we are aware, on its diversity related to geographic distribution. Hence, in the current study we investigated genotypic diversity of C. tropicalis especially in relation to their geographic origin from diverse regions of the globe. In addition, taxonomic features of this fungus were also investigated in an attempt to assess the existence of camouflaged sub species within the species. RAPD analyses of genomic DNA is a versatile tool for epidemiological studies of Candida species.[20] We have in previous studies used this technique to evaluate genomic diversity and clinical epidemiology of C. krusei ,[16] as well as C. albicans strains isolated longitudinally from HIV-infected individuals.[14] In the current study, a total of 36 clinical isolates of C. tropicalis were subjected to RAPD analyses and the fingerprinting patterns so generated were further evaluated using a well proven computer-assisted data analysis system. In general varying degrees of intra-species genomic similarity was observed confirming and extending previous reports on discriminatory power of DNA-based methods for strain delineation of Candida . The primers RSD8 and RSD9 were used with nondescript properties as they have been successfully tailored to analyse the yeast genome.[21] The rationale for using RSD12 was its previous utility in the analyses of the V3 variable region of the large ribosomal unit of C. albicans.[21] T3B has been employed to analyze the tDNA spacer region as this region is believed to be species-specific and variable amongst the different species of Candida .[22] In the current study, the primer RSD12 was used primarily to analyze the identical as well as unrelated isolates which were elicited by primer T3B. The data derived from most of the primers used indicated that isolates from some geographical locales could either be identical or disparate. A good example is the profile of Tanzanian isolates (TAN1 & TAN3) and Hong Kong isolates (T4, T6, T7, T9) that were identical at least with two of the primers used. However, the isolate TAN2 was unrelated to its counterparts (SAB of 0.62). Similarly, T1, T3 and HK2 were unrelated to their counterparts from Hong Kong. As all these isolates were obtained from individuals from the same geographic locale, it appears that strain diversity exists even within the same region. This has been previously shown for C. albicans in a number of studies.[4],[11],[12] It has been postulated that variations in the genotype could strategically enhance the survival of an organism in the oral cavity and this may be the reason for observed phenomenon in C. tropicalis . Alternatively, C. tropicalis strain may have undergone "genetic shuffling" for the reason that are yet unknown. Interestingly, the isolates HK2 and CA5, which originated from Hong Kong and Canada, respectively, revealed identical RAPD profiles with all four primers and fell into a single cluster on dendrogram analysis. Such existence of identical C. tropicalis isolates [analyzed by restriction digested ( Sfi I and BssH II) genomic DNA with pulsed-field gel electrophoresis] in different locales has been previously described.[4] In the latter report from seven different U.S. medical centers in Iowa Oregon, Georgia, Texas, New York, Michigan and California, C. tropicalis isolates with identical profiles were found.[3] Despite such genomic homogeneity of a few geographically-unrelated isolates, another cluster of isolates from Hong Kong (T9, T7, T6, T4 and T2) from HIV-infected individuals remained identical in a dendrogram generated with primer T3B and they formed a genetically distinct cluster (SABs 0.95 or greater) with primers RSD9 and RSD12. The latter observation suggests that the genetic relatedness based on geographic localization may be stronger as one would obviously surmise. On further investigation of PCR fingerprints generated by the three primers (RSD9, RSD8 and T3B) and their corresponding dendrograms we noted that a group of isolates (CH10, CA5, HK2, TAN2, G8 and G7) elicited distinct profiles that were dissimilar to their counterparts. The idiosyncratic behavior of the latter isolates was further confirmed on analysis of dendrogram generated by fingerprints of primer RSD12. It was interesting that the primer T3B, that has been previously tailored to analyze the species-specific tDNA repeats, produced such distinctly variable patterns. Perhaps, this is not surprising as genetically heterogeneous subgroups are not uncommon within species belong to fungi imperfecti . For instance Lehmann et al ,[23] reported two genetically distinct groups within species Candida haemulonni . The phenotypes of Candida famata were found among strains of Candida guilliermondii and several species of Debaryomyces 33 and more recently, C. dubliniensis was found as a distinct species camouflaged within C. albicans. Hence, this group of isolates in our studies, though classified as C. tropicalis based on phenotypic properties (API system), could represent either a new species closely resembling C. tropicalis or a subspecies within the species. Thanos et al[22] observed that profiles generated by primers T3B were species-specific using 24 different Candida species, although there were minor variations in some isolates within a single species. Similar observation has been made recently by Correia et al[2] using 177 Candida clinical isolates from two medical institutions in Portugal. However, in the current study, tDNA directed PCR fingerprints with the same primer revealed three distinct DNA profiles for both geographically-related as well as unrelated isolates. This, notwithstanding the vast majority (36 isolates) revealed either identical or similar band profiles confirming the observations of Thanos et al.[22] These observations tend to suggest that tDNA region in C. tropicalis is relatively homogeneous, yet distinct and the inverted repeats that are complimentary to T3B is either single or multiple. Most of the isolates that generated incongruent fingerprints with the primer T3B were those which produced idiosyncratic profiles with other primers. It is tempting to speculate that these isolates may represent subspecies within C. tropicalis. However, further work has to be conducted either to refute or confirm this hyphothesis. The current results clearly demonstrate that C. tropicalis is amenable to RAPD, although, different primers have varying powers of resolution, specificity and discrimination. The primer T3B elicited 14 genotypes (C1-C14) for 36 isolates and majority was classifiable as identical or non-identical. Hence T3B appear useful for identifying unrelated isolates, but not for highly-related and moderately-related isolates. On the other hand primers, RSD9 and RSD8 were much more discriminatory and elicited 29 (A1-A29) and 28 (B1-B28) genotypes, respectively and, identical isolates that were defined by primer T3B were split into highly and moderately related subclusters. Therefore, the latter two primers (i.e., RSD8, RSD9) appear to be more effective in grouping highly and moderately related isolates and perhaps to study the microevolution amongst C. tropicalis isolates. Further, rDNA based fingerprinting elicited more genotypes for identical types identified by the primer T3B, suggesting rDNA is genetically heterogeneous, but not distinct in C. tropicalis . Finally, on analysis of the mean SAB s value of RAPD profiles of isolates [Table - 4] from HIV-infected individuals, we were unable to show significant variations as they demonstrated similar diversity as isolates from healthy individuals. Diaz-Guerra et al[24] have previously attempted to study the genetic diversity of oral C. albicans isolates from HIV infected and healthy individuals from different geographical regions using a number of methods including PCR using phage core M13 sequence. They found, as in the current study, great deal of strain variation amongst C. albicans isolates irrespective of the health of the host. In conclusion, a fair degree of genetic similarity and diversity was found among C. tropicalis isolates from five different geographic regions. Contrary to expectations, identical genotypes from the same geographic locale were relatively limited, whilst significant genetic diversity between different regions was found only amongst one group of isolates from Scotland and, other regions. To our knowledge, this is the first report on the genetic profiles of oral C. tropicalis isolates from diverse geographic regions and our data provide interesting clues to the possible evolutionary trends within this Candida species in different regions of the world. References
Copyright 2006 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb06056f3.jpg] [mb06056t3.jpg] [mb06056t2.jpg] [mb06056t1.jpg] [mb06056f4.jpg] [mb06056t4.jpg] [mb06056f1.jpg] [mb06056f2.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}