 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
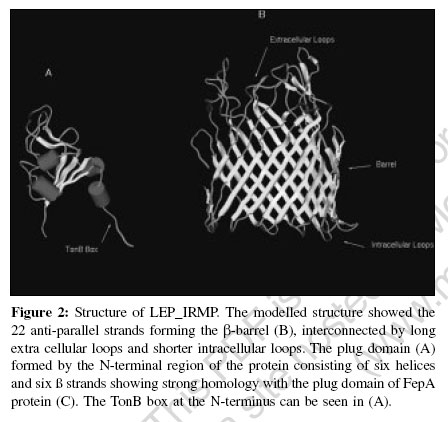
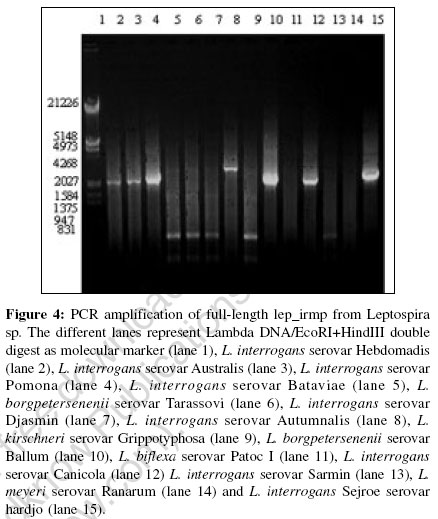
Indian Journal of Medical Microbiology, Vol. 24, No. 4, October-December, 2006, pp. 311-316 Supplement Understanding iron acquisition by pathogenic leptospires: A review Sritharan M, Asuthkar S, Sridhar V Department of Animal Sciences, School of Life Sciences, University of Hyderabad, Gachibowli, Hyderabad - 500 046, Andhra Pradesh Code Number: mb06099 Biochemical role and bioavailability of iron Iron is the second most abundant metal after aluminum and the fourth most abundant element in the earth′s crust. Iron is involved in many important metabolic functions of plants, animals and microorganisms. Iron, by virtue of its ability to accept and donate an electron is important in biological systems in terms of catalysis of several biochemical reactions and transport of reducing equivalents in the electron transport chain. Fe 2+ Fe 3++ e - However, its insolubility at biological pH makes it unavailable, as it exists as insoluble ferric hydroxides and oxyhydroxides. Nature perhaps limited the amount of free iron due to its toxicity in the aerobic environment, as it catalyses the formation of extremely reactive free radicals in the presence of oxygen and triggers chain reactions that are harmful to biomolecules, mainly DNA and proteins. Microorganisms are no exception to the need for iron and in the aerobic environment iron exists primarily in the oxidized, ferric form (Fe 3+), which at pH 7.0 has a solubility of 1.4 x 10 -9M 1 that is too low to support the growth of microorganisms. Consequently, microorganisms have evolved complex systems to efficiently capture iron and regulate its acquisition. Various means of solubilizing iron and then holding it in an utilizable form have evolved in both pathogens and non-pathogens. The acquisition of iron by a pathogen is recognized as one of the contributing factors in the successful establishment of the pathogen within the host. In the host-pathogen interaction, the balance between the ability of a mammalian host to withhold iron from invading micro-organisms and the ease with which the micro-organism can acquire this iron from the host is critical. Withholding of iron[2] is one of the non-specific mechanisms of defense that include other mechanisms like physical barriers, bacterial inactivation, phagocytosis with intracellular killing, elaboration of natural antibodies etc. Iron is held as protein-bound iron by transferrin, lactoferrin and ferritin that ensure that very little free iron is found in the circulation. Having two similar, but not identical, binding sites for iron characterize transferrin and lactoferrin found extracellularly. Both sites have binding constants for iron of ~10 20. Ferritins are the primary iron storage compounds of most organisms with a molecular weight of ~ 5 x 10 5 composed of 24 protein subunits that form a hollow sphere that can accommodate> 4000 atoms of Fe 3+ as a macro-organic complex. Microbial acquisition of iron Microorganisms have adapted to conditions of low iron by the elaboration of novel iron acquisition machineries. The more commonly found mechanism is the expression of low molecular weight Fe 3+sub - specific ligands called siderophores and their receptors, the iron related/regulated proteins (IRPs/IREPs). The other equally important strategy adopted specifically by pathogenic bacteria is the direct removal of the protein -bound iron by specific receptors for the host iron-containing molecules like transferrin, lactoferrin, hemin and hemoglobin. There are extensive reviews on the iron-acquisition machinery in several bacteria.[3],[4],[5],[6],[7],[8],[9] a) Siderophore mediated iron acquisition The expression of siderophores and the ferric-siderophore receptors, the iron- regulated membrane proteins/ envelope proteins (IRMPs/IREPs) is the most common and well-studied mechanism of iron acquisition. The affinity of the siderophore for Fe 3+is very high with association constant (Ks) ranging from 10 22to 10 50that renders it capable of removing iron from the host iron-binding proteins. Siderophores have been demonstrated in several bacteria including Yersinia spp (yersiniabactin), Pseudomonas aeruginosa (pyochelin), Vibrio spp. (vibriobactin), Escherichia coli (enterobactin and aerobactin), Mycobacterium spp. (intracellular mycobactins and extra cellular carboxymycobactins/exochelins)[7] etc. TonB- dependant ferric-siderophore receptors Uptake of iron (Fe 3+) bound to siderophores, heme and host proteins (transferrin and lactoferrin) is mediated by specific outer membrane (OM) receptors.[10] These receptor-mediated high-affinity iron transport systems utilize the cytoplasmic membrane proton-motive force as an energy source that depend crucially on the TonB protein working in concert with ExbB and ExbD. The TonB protein is anchored in the cytoplasmic membrane by an uncleaved leader sequence and spans the periplasmic space to interact directly with the outer membrane components. The conformational change in this protein mediates the coupling of the cytoplasmic membrane energy to the high-affinity active transport of heme and Fe 3+ -siderophore complexes across the outer membrane. The subsequent transport across the cytoplasmic membrane to the inside is carried out by an ABC transport system. Many of these iron transport receptors are multi-functional and mediate the transport of vitamin B12 and certain colicins.[10] The identification of the ferric-siderophore receptors from E. coli , namely the FepA receptor for ferric enterobactin, FecA receptor for ferric citrate and FhuA receptor for ferrichrome led to studies in other gram-negative bacteria and the identification of a number of TonB-dependant OM ferric-siderophore receptors.[9] All of them are expressed at high levels only under iron-deficient conditions. All these OM siderophore receptors are related[11] and the crystal structures of FepA, FecA and FhuA have been determined.[11],[12],[13] These proteins consist of a large β -barrel domain, consisting of 22 antiparallel β -strands that traverse the OM allowing the siderophore complex to access the periplasm. Part of this domain protrudes to the exterior as the extracellular loop region and contains residues for binding to the specific ligand; it is thus not surprising that FepA, FecA and FhuA differ in these extra cellular loops, thus explaining the specificity of the respective receptor for its ligand despite the overall similarity in their conformation and mechanism of transport of the ferri-siderophore complexes. This β -barrel tube is blocked by a plug unlike the porins. The ′plug′ or ′cork′ consists of a globular domain derived from ~160 residues at the N-terminus and is positioned in the pore towards the periplasmic end of the barrel. The plug gates the channel (external diameter of ~4 nm) limiting access to the periplasm. Mechanism of iron uptake: Sequence of events upon binding of ferric-siderophore by its receptor Though the binding of ferri-siderophores to OM receptors does not require any specific factors, transport of ferri-siderophores through OM receptors requires energy. This energy is provided by the proton motive force in the cytoplasmic membrane generated by the energy-transducing TonB-ExbB-ExbD protein complex.[14] This system, involving the energy transduction process, well studied in E. coli requires direct contact between TonB and OM receptors. A conserved hydrophobic seven-amino acid segment, the ′TonB box′, at the N-terminus of TonB-dependent OM receptors has been found to be required for TonB-mediated uptake of ferri-siderophores.[14] Upon binding of the ferric siderophore to the receptor, a conformational change, mediated by TonB causes the translocation of the associated ferri-siderophore to the periplasm. The barrel domain, kept closed by the plug opens by subtle rearrangements of the loops of the plug domain, thus allowing the formation of a small channel through which the siderophores are translocated. The transport of siderophores into the cytoplasm across the periplasm and cytoplasmic membrane is facilitated by the periplasmic binding proteins and the cytoplasmic ABC permeases. The periplasmic binding proteins consist of the FecB protein of the Fe 3+-dicitrate transport systems, the FepB protein of the ferri-enterochelin transport system and FhuD protein of the ferric hydoxamate uptake systems. Although E. coli K-12 possesses six iron-transporting OM receptors, it only contains three associated binding-protein-dependent ABC systems: one each for the transport of ferric citrate, ferric hydroxamates and ferric catechols. Thus, the OM receptors display a greater specificity than the binding-protein-dependent ABC systems. Once the ferric-siderophore is internalized and is in the cytosol, the iron is presumed to be released by reduction of the ferric to ferrous form, by virtue of the relatively low affinity of the siderophore for ferrous iron. b) Direct uptake of iron from host proteins Uptake from transferrin and lactoferrin Pathogenic bacteria such as Neisseria spp. elaborate independent transferrin (Tf) and lactoferrin (Lf) receptors, that are OM receptors induced upon iron limitation.[8] Like the ferric-siderophore receptors, these receptors are also TonB-dependant. However, the iron is released at the cell surface itself and internalized while the transferrin and the lactoferrin are released extra cellularly. These receptors are composed of two subunits: Tbp1 and Tbp2 for transferrin receptor and Lbp1 and Lbp2 for lactoferrin receptor. Tbp2 and Lbp2 are homologous but are not related to OM siderophore receptors.[15] They are lipoproteins anchored by their N-terminal lipid groups to the outer surface of the OM. Tbp1 and Lbp1 have affinity for transferrin and lactoferrin respectively and they appear to favour the binding of the receptor to their respective ligands and further to differentiate between the iron-loaded and iron-free proteins.[16] Unique periplasmic and ABC transporters mediate the internalization of the iron released from Tf and Lf across the periplasm. Uptake from hemin and hemoproteins Several pathogenic bacteria utilize heme as a source of iron; e.g., include Vibrio spp, Sheigella dysenteriae , Enterhemorrhagic E. coli 0157:H7, Y. enterocolitica , Y. pestis, Neisseria spp, S. marcescens, P. aeruginosa and H. influenzae .[3] Two common mechanisms are involved in the uptake of iron from heme and hemoproteins. The first and most common mechanism is the direct binding and processing of the heme or hemoprotein by a TonB-dependent, OM receptor and passage of the released heme to the periplasm for transport to the cytoplasm like the ferric-siderophores. The second involves a soluble protein or hemophore that interacts with the hemoproteins and returns heme to the cell surface, where it interacts with a TonB-dependent receptor. A large section of Ton B-dependent OM proteins from gram-negative bacteria have been identified and characterized as heme or hemoglobin-binding receptor proteins. All the bacterial heme uptake systems must first release the heme from its particular protein scaffold and transport it to the periplasm. Serratia marcescens elaborates an extra cellular protein, HasA, under iron limitation, which is necessary for hemoglobin to be used as an iron source. This hemophore does not directly affect the release of iron from hemoproteins but carries a heme-binding activity and can remove heme from hemoglobin.[16] Histidine residues within the heme receptor proteins may act as axial ligands of heme-iron and could play a major role in the internalization of heme. Iron levels and regulation of expression of components of the iron acquisition machinery Intracellular iron, in association with a regulatory protein modulates the expression of the components of the iron acquisition machinery.[7] In several Gram negative bacteria, the Fur (ferric uptake regulator) protein, identified in E. coli as a small 17 kDa molecule, binds to iron under conditions of iron sufficiency and this Fe 2+sub -Fur complex binds to the Fur box upstream of the genes involved in the synthesis of the components of iron acquisition, thereby blocking their transcription. The Fur box is a 19 bp degenerate palindromic sequence GATAATGAT(A/T)ATCATTATC, rich in A-T bases. The corresponding equivalent in Gram-positive bacteria is the 25 kDa DtxR repressor (so called, as it was first identified upstream of the tox gene encoding the diphtheria toxin in Corynebacterium diphtheriae ). Identification of a hemin-binding protein in pathogenic Leptospira spp. In silico analysis of the genome of Leptospira interrogans serovar Lai with ferric enterobactin receptor FepA of E. coli identified a putative TonB-dependent outer membrane receptor protein gene sequence (Gen Bank accession number LB191).[17] The translated product, referred to as LEP_IRMP (leptospiral iron-regulated membrane protein), despite showing low level of similarity (39%) and identity (22%) with FepA of E. coli , revealed features of protein folding like other Fe 3+sub -siderophore receptors [Figure - 1]a and b. Upon homology modeling with Insight II, using FepA as the reference protein, the 3D structure of LEP_IRMP was found to show the characteristic β -barrel structure with the three domains, namely the β -barrel, plug domain and N-terminal TonB box [Figure - 2]. The high degree of structural similarity between LEP_IRMP and FepA is evident. The β -barrel domain, forming the trans-membrane part of the receptor consisted of the 22 anti-parallel strands with large loops extending towards the extra cellular side and short periplasmic turns facing the periplasmic side. As expected, there were striking differences in the sequence between LEP_IRMP and FepA in the extra cellular loops, reflecting their ligand specificity. As indicated above, the FepA, FhuA and FecA receptors of E. coli also differ in these sequences reflecting ligand specificity to ferric enterobactin, ferrichrome and ferric citrate respectively. The plug domain of LEP_IRMP, consisting of six helices and six β strands in the N-terminal region shares a high degree of homology with FepA protein. The TonB box in LEP_IRMP, represented by the sequence ESQIVVTGS was located between amino acids 79-87 and it differed from the consensus sequence in the third amino acid Q. BLASTP analysis of microbial genomes showed that LEP_IRMP shared strong homology with ferric siderophore protein receptors/heme-binding proteins/ colicin receptors/ Vitamin B12 receptors of several Gram-negative bacteria; this is not an unexpected observation, as iron transport systems are known to facilitate the transport of colicins and vitamin B12 in several Gram-negative bacteria.[18] In addition to the strong structural similarity of LEP_IRMP with other ferric-siderophore receptors, the presence of the fur gene (LB183) and the Fur box in the vicinity of LB191 (encoding LEP_IRMP) [Figure - 3] confirmed that this is an iron-regulated protein.[17] LEP_IRMP is a putative hemin-binding protein. This was confirmed by bioinformatics tools and also verified experimentally. The gene encoding heme oxygenase (LB186) was located in the neighborhood of lep_irmp and the FRAP-NPNL motif, associated with heme-binding was also seen in LEP_IRMP. Experimental proof (results under communication) also pointed to this protein as a hemin-binding protein. Further, we could not detect any siderophores in the culture filtrate of pathogenic leptospires. Interestingly, we found that the full-length 2148 bp lep_irmp gene [Figure - 4] is seen in several pathogenic serovars[17] including L. interrogans serovars Hebdomadis, Australis, Pomona, Canicola and Hardjo Autumnalis and L. borgpetersenii serovar Ballum and is absent in non-pathogenic serovars. In some of the pathogenic serovars including L. borgpetersenii serovar Tarassovi, Djasmin, L. kirschneri serovar Grippotyphosa, L. weilii serovar Sarmin , we identified smaller size products . These serovars were also negative for the 285 bp product by PCR with G1/G2 primers.[19] It is quite likely that Leptospira , like other bacteria express more than one iron transport system. Whether the expression of LEP_IRMP by some serovars is associated with the adaptation of these serovars to particular animal hosts remains to be answered. Virulent Leptospira strains, but not avirulent or saprophytic strains, have been shown to exhibit chemotaxis towards haemoglobin.[20] Many pathogenic bacteria can obtain iron from cell-free haemoglobin or heme for multiplication in-vivo .[21] Further, it is the iron in these molecules that appear to be important.[22] As mentioned above, heme is one of the most effective nutritional source of iron for several pathogenic bacteria. Heme-binding and bacterial virulence is reported in several bacteria.[21] To gain access to the intracellular heme, pathogenic bacteria secrete hemolysins.[21]It is well-established that Leptospira elaborate several hemolysins and hemolysin-like activities.[23] Our lab, also focusing on leptospiral hemolysins has found evidence for the association of iron with hemolysin expression (under preparation). Detailed analysis of the association between iron, expression of iron-regulated membrane proteins and hemolysins in pathogenic leptospires will help in the better understanding of host-pathogen interrelationship and aid in the identification of potential drug targets. Conclusion Pathogenic Leptospira spp, like several other pathogenic bacteria express novel iron-regulated proteins that are probably not only involved in the acquisition of this essential nutrient, but is also associated with virulence. As hemolysis, hemorrhage and jaundice are encountered in several cases of leptospirosis, a better understanding of the role of this element in the pathogenesis will help in the development of methods for the control of this disease that is of considerable concern in our country with the recurring floods and improper civic amenities prevailing in several parts of the country. Acknowledgements The authors wish to thank Andhra Pradesh -Netherlands Biotechnology Programme for the establishment of containment facilities and Department of Biotechnology for funding part of the work on iron-regulated proteins in leptospires. Sridhar V acknowledges CSIR for providing SRF and Swapna A acknowledges ILS-UoH fellowship. References
Copyright 2006 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb06099f3.jpg] [mb06099f1.jpg] [mb06099f4.jpg] [mb06099f2.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}