 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
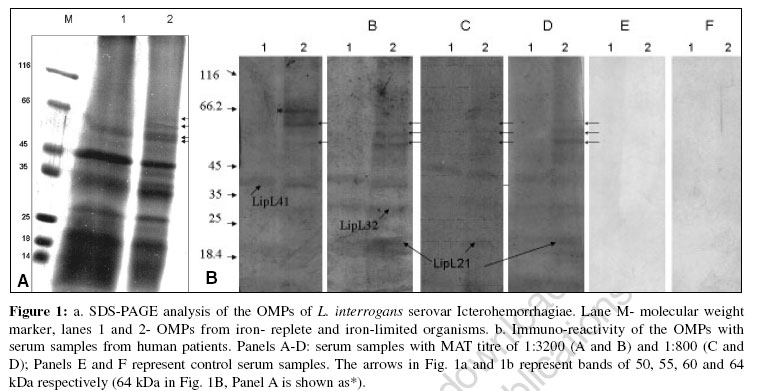
Indian Journal of Medical Microbiology, Vol. 24, No. 4, October-December, 2006, pp. 339-342 Supplement Iron limitation and expression of immunoreactive outer membrane proteins in Leptospira interrogans serovar icterohaemorrhagiae strain lai Velineni S, Asuthkar S, Sritharan M Department of Animal Sciences, School of Life Sciences, University of Hyderabad, Hyderabad - 500 046, Andhra Pradesh Code Number: mb06107 Iron (Fe) is an important element, essential to the growth of almost all living cells. It is needed for important cellular functions such as transport and storage of oxygen and plays an important role in the transfer of electrons in the electron transport chain. Since it has an extremely wide redox potential, it is a very versatile biocatalyst. A variety of enzymes such as catalase, peroxidase and dehydrogenase require iron for activity. It serves as a cofactor in key metabolic processes such as nucleotide biosynthesis and energy production.[1] Iron, by virtue of its chemical nature does not exist in a freely available soluble form and exists as the insoluble ferric hydroxide inside the host. In the mammalian host, the iron that is found extracellularly is bound to carrier proteins called "siderophilins". The principal siderophilins in the blood are haemoglobin and transferrin, while lactoferrin predominates in the secretions at mucosal surface. Microorganisms have adapted to this low iron environment by the induction of novel pathways for obtaining iron. Acquisition of iron is one of the factors contributing to the virulence of a pathogen. Most bacteria respond to limiting iron conditions prevailing within the mammalian host by the production of siderophores and specific receptors on the cell surface, the iron-regulated proteins (IRPs).[2],[3] Some pathogenic bacteria obtain the iron directly by elaborating specific receptors on their cell surface for host iron-binding proteins, namely transferrin, lactoferrin and hemin as seen in Neisseriae spp.[4] At the molecular level, iron controls the expression of virulence factor(s) like toxins, hemolysins in many microorganisms like Corynebacterium diphtheriae, Escherichia coli, Pseudomonas aeruginosa and other microorganisms.[5] Leptospirosis is a spirochetal disease caused by the pathogenic members of the genus Leptospira . It is a wide-spread zoonosis that affects humans worldwide, in both urban and rural areas and in temperate and tropical climates.[6],[7] Humans are accidental hosts and are infected by contact with an environment contaminated by urine of shedder hosts such as rodents, cattle and dogs. Individuals at risk include farmers, abattoir workers, sewer workers and miners as all occupations related to animal holdings are at a great risk. The clinical spectrum of the disease ranges from mild flu-like illness to severe fatal form due to multi-organ failure conventionally called as "Weil′s disease". The clinical presentation varies from patient to patient which includes hepato-renal failure, myocarditis, severe pulmonary hemorrhage with respiratory distress and meningitis.[8] Leptospirosis at present is greatly under-reported due to lack of simple, rapid and efficient tests for early diagnosis and also vaccines that can elicit protective immunity. Serological techniques for detection of antibody, isolation and demonstration of leptospires and polymerase chain reaction (PCR) are available laboratory methods for diagnosis. Low success rate of isolation, unreliability of direct demonstration of leptospires using dark field microscopy and inaccessibility of molecular techniques like PCR-based diagnosis to most peripheral hospitals and clinics make serological tests play an important role in laboratory diagnosis of leptospires. Though the microscopic agglutination test (MAT) continues to remain the "gold standard" for diagnosis, it has its limitations. The immense serovar diversity among the pathogenic leptospires necessitates maintenance of a large panel of serovars, thus restricting the use of the test. The test is also complex to perform and interpret. The lack of good and user friendly diagnostic test has led to a plethora of tests being developed which includes latex agglutination (LA), haemagglutination assays (HA), LeptoTek Dri-Dot, Lepto dipstick, Lepto lateral flow, microcapsule agglutination test (MCAT), enzyme-linked immunosorbent assay (ELISA) and dot-ELISA have been evaluated as rapid screening tests for leptospirosis. Most of these techniques were found to be more sensitive during the acute stage of the disease.[9] Much work has focused on the role of leptospiral lipopolysaccharides (LPS) in immunity[10] and the genetics of LPS biosynthesis have been partly elucidated.[11] Preparations of leptospiral LPS can elicit protective immunity. These currently available leptospiral LPS vaccines have low efficacy, are serovar specific and generally produce only short-term immunity in domestic livestock. Efforts to identify immunogenic components of value in diagnosis and in vaccine preparation resulted in characterization of LipL 21, LipL 32, LipL 36, LipL 41 and LipL 48 outer membrane proteins.[12],[13],[14] Recently, it was reported that by screening with immune sera from leptospirosis patient, 16 proteins were identified as potential vaccine candidates or to be used in diagnosis. Out of these sixteen, four proteins such as OMPAL 21, LipL 23, OMPL 31 and MPL 36 were not expressed in L. biflexa and conserved among pathogenic leptospires.[15] It was reported that two proteins (31 and 41 KDa) function synergistically in the immunoprotection of hamsters suggesting that an effective protein-based vaccine would contain several components.[16] The search for protective immunogen and candidate antigen for diagnosis is complicated by the possibility that the expression of some of these important components may be influenced by conditions prevailing within the immediate environment of the pathogen within the mammalian host. As iron limitation is one of the non-specific mechanisms of the host immunity, we studied the influence of expression of iron limitation on the outer membrane proteins in L. interrogans serovar Icterohemorrhagiae strain Lai and we report the immunoreactivity of serum from human patients with leptospirosis with specific outer membrane proteins expressed under conditions of iron deprivation and also the effect of iron deprivation on the expression of LipL32 and LipL41. Materials and Methods Growth conditions Leptospira interrogans serovar Icterohaemorrhagiae strain Lai (obtained from the Regional Medical Research Centre and WHO collaborating center for Diagnosis, Reference, Research and training in leptospirosis (ICMR), Port Blair Andaman and Nicobar Islands) was maintained in semisolid (0.15% Noble agar) EMJH supplemented with 10% enrichment (Difco, USA) at 30°C in screw-capped test tubes. Iron deprivation was established as follows. Iron-free glassware and glass double-distilled water was used. Initially, the organisms were cultured in EMJH liquid medium supplemented with 10% enrichment and after sufficient growth, the 150 uM 2-2′ dipyridyl was added gradually over a period of two days to gradually deprive them of iron. These cells were then re-suspended overnight in iron-free medium that was pre-incubated overnight with 150 uM 2-2′ dipyridyl. An identical set of culture without chelator representing iron-replete organisms was maintained. All the cultures were harvested and analyzed for the expression of iron-regulated membrane proteins. Extraction of outer membrane proteins by Triton X-114 Triton X-114 extraction was done based on the method of Haake et al .[17] The bacterial cells were harvested by centrifugation at 10,000 rpm for 20 min and washed thrice with 50 mM Tris (pH 8.0) + 5 mM MgCl 2 and then incubated overnight at 4°C in 2% Triton X-114 in 10 mM Tris (pH 8.0) + 1 mM EDTA + 150 mM NaCl. The insoluble material (cell pellet) was separated by centrifugation at 17, 000 x g for 10 min and supernatant was added with 20 mM CaCl 2 . Phase separation was performed by warming the supernatant to 37°C and subjecting it to centrifugation at 2000 x g for 15 min. The proteins in the detergent phase (containing outer membrane proteins) were precipitated with acetone, solubilised in Tris-HCl buffer with 0.6% SDS, protein estimated by BCA method and subjected to SDS-PAGE. Electrophoretic separation by SDS-PAGE The outer membrane protein sample was boiled for 10 min in the sample buffer (60 mM Tris- HCl pH 6.8 + 20% glycerol + 0.6% SDS + 0.1% bromophenol blue) and then subjected to discontinuous SDS-PAGE[18] on a 5-20% gradient gel. The bands were visualized after staining with Coomassie Brilliant blue. Serum samples Serum samples from human patients who tested positive by LeptoTek DriDot test for leptospirosis were obtained from the Department of Microbiology at NIMS, Hyderabad. MAT was done by routine procedures on these serum samples, which were aliquotted and stored at -20°C. Specific antibodies against LipL32 and LipL41 were kindly provided by David Haake (University of California, USA). Immunoblotting analysis Proteins were transferred onto nitrocellulose membranes with a Trans-Blot electrophoretic transfer cell using Towbin′s buffer.[19] The membranes were initially blocked with 5% non-fatty milk powder in Tris- buffered saline with 0.05% Tween 20 (TTBS) for 2 hrs and washed thrice with TTBS and incubated with a 1/600 dilution of human patient′s sera with healthy controls, 1/10,000 dilution of anti-LipL32 serum and 1/8000 dilution of anti-LipL41 serum separately. Binding of antibodies was detected by using 1/500 dilution of ALP-conjugated rabbit anti-human or goat anti-rabbit immunoglobulin G. Immunoblots were developed by using the BCIP-NBT detection system (Bangalore - Genei, India). Results and Discussion Establishment of conditions of iron limitation has not been easy with leptospires as they are very sensitive to the presence of iron chelators and also to low iron levels (as was seen in cultures to which no iron was added). When culture medium was freshly inoculated for growth under iron-deficient conditions, growth was significantly lowered and soon death and disintegration of the cells were seen. We thus established sufficient growth in the regular media and then induced iron limitation gradually by the addition of iron chelators. In the last stage, the medium was removed by centrifugation and the cells were re-suspended in medium that was made iron-free by incubating the medium overnight with the iron chelator. This was necessary for the induction of expression of iron-regulated proteins. OMPs of L. interrogans serovar Icterohemorrhagiae were separated electrophoretically and subjected to immuno-blotting with four serum samples positive by MAT for L. interrogans serovar Icterohemorrhagiae and two control serum samples. In addition to the reactivity of the immunodominant LipL41, LipL32 and LipL21 in all the samples [Figure - 1]B, as against the control which showed no reactivity with any leptospiral protein, we observed four immuno-reactive OMPs of approximate molecular weights 50, 55, 60 and 64 kDa respectively from leptospires grown under conditions of iron limitation [Figure - 1]A, lane 2 and [Figure - 1]B Panels A-F, lane 2, indicated by arrows). The former three were detected by all the sera while the 64 kDa band was found to react with one of the serum samples ([Figure - 1]b- Panel A, lane 2). The relevance of the control of expression of these proteins by iron and their immunogenicity in-vivo needs further studies to understand their role in host-pathogen inter-relationship. Iron deprivation and the expression of virulence determinants is a well-studied aspect of host-pathogen interrelationship in several microorganisms.[5] Pathogenic leptospires also express hemolysins and sphingomyelinases. It would be worth analyzing if one or more of these proteins serve as virulence determinants. When OMPs from iron-replete and iron-limited organisms were treated with antibodies against recombinant LipL41 and LipL32 respectively, anti-LipL41 showed significant reactivity with Lip41 as anticipated [Figure - 2]a; however, it also reacted with a 32 kDa band only in iron-replete organisms (lane 1) with no reactivity in the corresponding iron-limited cells. Anti-LipL32 reacted with the 32 kDa band in OMPs of both iron-replete and iron-deficient organisms, with the latter showing greater reactivity. This antibody also reacted with the LipL41 ([Figure - 2]A, lanes 1 and 2). Though the role of iron in the expression of some of the OMPs is evident, the significance of these observations, especially with reference to the LipL41 and LipL32 needs further studies to deduce any conclusions. A better understanding of the host-pathogen interrelationship with reference to iron will aid in the identification of candidate antigens for diagnosis and development of drug targets for the control of this disease. Acknowledgements Velineni Sridhar acknowledges the Senior Research Fellowship given by CSIR. Asuthkar Swapna acknowledges the Senior Research Fellowship by ILS-MoU scheme of the University of Hyderabad. Manjula Sritharan acknowledges the AP-Netherlands Biotechnology Programme and Department of Biotechnology for the establishment of containment facility and research in leptospirosis. References
Copyright 2006 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb06107f2.jpg] [mb06107f1.jpg] |
| |||||||||
{kind=link}
{kind=link}