 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
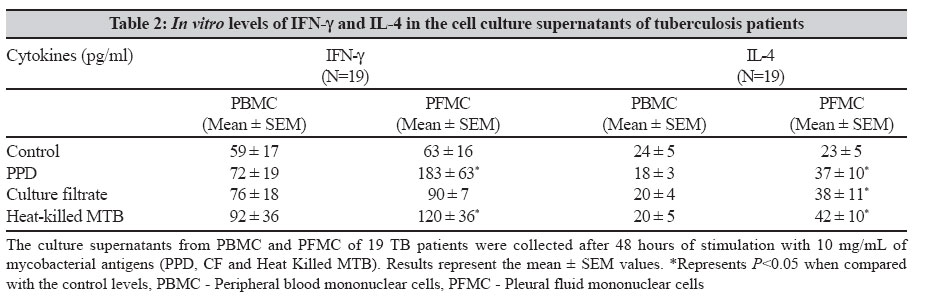
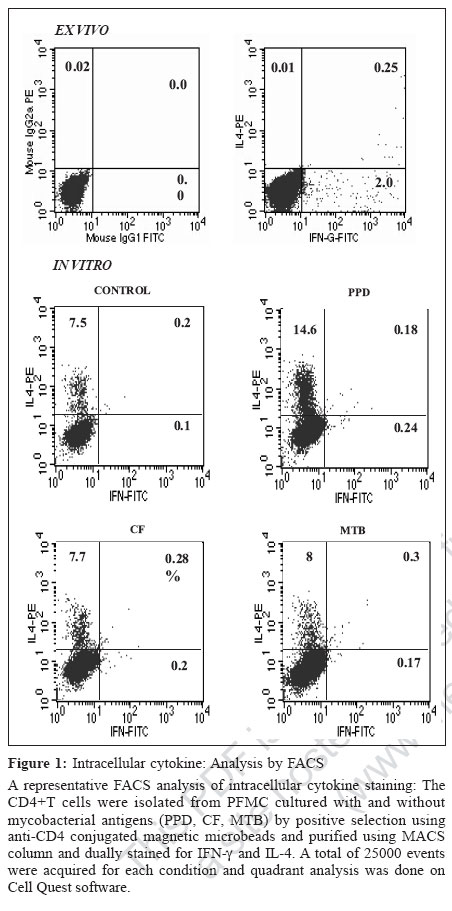
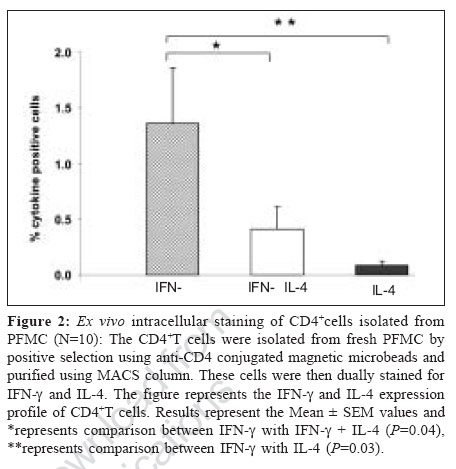
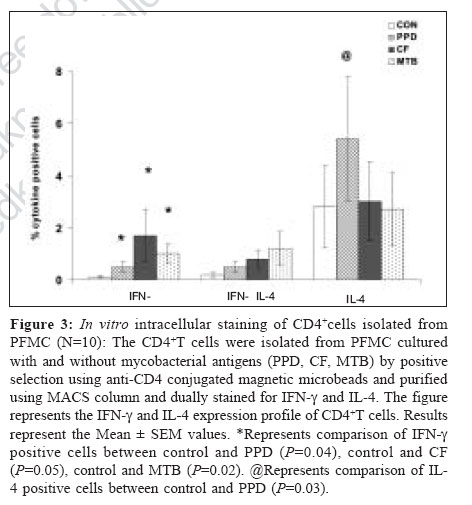
Indian Journal of Medical Microbiology, Vol. 25, No. 1, January-March, 2007, pp. 18-23 Original Article Differential T helper cell response in tuberculous pleuritis Prabha C, Jalapathy KV, Matsa RP, Das SD Department of Immunology, Tuberculosis Research Centre (ICMR), Chennai - 600 031, Tamilnadu Date of Submission: 23-Jun-2006 Code Number: mb07006 Abstract Purpose: This study was conducted to understand the in vivo and in vitro immune responses and to find whether there exists any difference in the systemic and localized immune responses in tuberculous pleuritis.Methods: The in vivo levels of IFN-γ and IL-4 were compared in plasma (BL) and pleural fluid (PF) of 47 tuberculous (TB) and 31 nontuberculous pleuritis (Non-TB) patients. In vitro cytokine response to various mycobacterial antigens was studied in 19 TB patients by ELISA . Both ex vivo and in vitro cytokine responses were further ascertained by intracellular cytokine staining on purified CD4+ T cells from pleural fluid mononuclear cells (PFMC) of 10 TB patients. Results: The ex vivo results showed a significant increase in IFN-γ levels and higher IFN-γ + T cells in PF. On the other hand, in vitro results showed simultaneous increase in both IFN-γ and IL-4 levels in the supernatants of antigen stimulated PFMC. Similarly antigen specific increase was observed in both IFN-γ + and IL-4+ T cells in all cultured conditions. However, the percentile increase was more in IL-4 secreting T cells, significant for PPD stimulation ( P <0.05), indicating that in vitro cellular response was dominated by Th2 type. Conclusions: These results showed a differential T-helper response in TB pleuritis suggestive of predominant Th1 in vivo and mixed response (Th1 and Th2) under in vitro conditions. Keywords: Cytokines, immune response, pleuritis, t cells, tuberculosis Antigen recognition through T-cell receptors results in the production of cytokines that amplify a specific response required for antigen elimination. These cytokines mediate and regulate the cell-cell communication between the antigen presenting cells (APCs), lymphocytes and other host immune cells in the course of an immune response. Protective or immuno-pathologic outcome depends on the production of appropriate set of cytokines in response to an infectious agent. Inhibition strategies adapted by the infectious agents against the protective cytokine response leads to the severity of the disease either by up regulating the antagonistic cytokine response or by inducing mechanisms that lead to the depletion of the protective cytokine response. Cytokines secreted by T helper (Th) cells against the infectious agents are of critical importance for the outcome of many diseases. T helper cell response can be divided into Th1 or Th2 based on the pattern of cytokine secretion. Th1 cells produce gamma interferon (IFN-γ), Interleukin-2 (IL-2), Tumor necrosis factor-β (TNF-β ) whereas Th2 cells produce IL-4 or IL-5.[1] T-cells that produce cytokine pattern distinct from the well-defined Th1/Th2 sets have been described and named as Th0, Ths or Tr.[2],[3] Th2 cells offer protection against the helminthes infections whereas Th1 cells are essential for protection against intracellular bacteria.[4] In infection with M. tuberculosis, a Th1 response has been shown to be protective.[5] Interferon gamma (IFN-γ), a Th1 type cytokine is essential in tuberculosis immunity, which is the single most important factor for macrophage activation.[6] Mice, which lack either IFN-γ or the IFN-γR, are highly susceptible to mycobacteria that show its importance for the control of tuberculosis.[7] The pro-inflammatory response initiated by Th1 cytokines is antagonized by anti-inflammatory cytokines. IL-4 is a prototype anti-inflammatory cytokine that induces Th2 phenotype development and prevents overshooting of Th1 response at the systemic level. It cross regulates IFN-γ production indirectly by inhibiting its responsiveness to IL-12. Although some reports have shown that IL-4 is not essential for protective immune response against mycobacteria,[8],[9] the report by Sugawara et al showed that IL-4 is required for defense against mycobacterial infection. However the exact role of this cytokine remains to be established in tuberculosis.[10] There are many reports based on either the in vivo levels or on the in vitro studies, yielding different results on the pattern of T-cell cytokines and thus T-helper cell response to tuberculosis.[9],[11],[12] We conducted this study to evaluate the type of immune response (Th1/Th2) in vivo and in vitro in concert with the systemic and localized response. We have recruited tuberculous pleuritis patients for our study as they provide a model to understand immuno protective role due to self-limitedness of the disease. Moreover the pleural fluid cells that closely encounter tuberculous bacilli and generate appropriate immune response against tuberculosis, are a better source to study the immunity at the site of disease. Our results showed that there is a differential T-helper cell response at the periphery and the local site and under in vivo and in vitro conditions in tuberculous pleuritis. Materials and Methods Patient population During the study period of two years (2002-04), 78 patients with pleural effusions were recruited from the Government General Hospital, Chennai. The patients were grouped into two, based on various diagnostic criteria. TB group consisted of 47 patients with effusion due to tuberculosis infection. Non-TB group consisted of 31 patients with transudative effusions due to congestive cardiac failure and liver disorders. We expected 50% mean difference between the two parameters. To pick up the difference, minimum sample required with 80% power was 34. Hence, the sample size of 47 TB and 31 Non-TB patients was adequate. The diagnosis for tuberculosis was on the basis of any two positives with the following criteria: (i) Smear positivity for sputum/pleural fluid for M. tuberculosis (Ziehl-Neelsen method), (ii) Culture positivity for the growth of M. tuberculosis on Lowenstein-Jensen medium, (iii) Clinical picture that included chest X-ray and clinical symptoms for tuberculosis and (iv) PCR positivity for the presence of the insertion sequence IS6110 specific for M. tuberculosis. The age of these patients varied from 20-61 years. These patients were found to be seronegative for HIV. The samples were collected before the start of the treatment. A written informed consent was obtained from each patient. The sample collection and the study followed the ethical guidelines of Government General Hospital, Chennai. Mononuclear cell preparation Peripheral blood mononuclear cells (PBMC) and pleural fluid mononuclear cells (PFMC) were isolated from blood and pleural fluid by Ficoll-Hypaque density gradient centrifugation at 1800 rpm for 30 minutes. The cells were then washed with HBSS (Whittaker) and RPMI 1640 (Sigma Chemical Co, St. Louis, Mo) at 1500 rpm for 10 minutes. The PBMC and PFMC were constituted in RPMI 1640 at the ratio of 1 x 10 6 cells/mL. The medium was supplemented with 10% autologous serum and 2 mM L-glutamine (Sigma Chemical Co). The viability of the cells was assessed by trypan blue exclusion method. Stimulation of mononuclear cells Nineteen out of 47 patients from TB group, who were positive by all the diagnostic criteria and had adequate cell numbers in their pleural fluid, were included for the in vitro studies. The PBMC and PFMC at the density of 1C10 6 cells/mL were cultured in 48 well plates (Costar, Cambridge, Mass) with or without antigens. The cells cultured in the absence of M. tuberculosis antigen served as control. The antigens used for the stimulation were purified protein derivative (PPD, Weybridge, U. K.), culture filtrate (CF) and heat killed M. tuberculosis (MTB) at the concentration of 10 mg/mL. The culture filtrate antigen was prepared in our laboratory by growing the standard M. tuberculosis H37Rv strain in Sauton′s medium for six weeks. The culture filtrate was concentrated and its protein content was estimated using protein estimation kit (Bangalore Genei) and diluted to required concentration. Heat killed preparation of H37Rv strain was obtained by killing the bacilli at 80°C for 30 minutes. The PBMC and PFMC were cultured in the presence of these antigens. The supernatants were collected after 48h, when the cytokine levels were found to be highest in initial standardization experiments. The cell-free supernatants were stored at -70°C for the cytokine assessment. Purification of CD4+cells The CD4+T cells were isolated from fresh PFMC and also from PFMC cultured with and without antigens by positive selection using anti-CD4 conjugated magnetic microbeads and purified using MACS column according to the manufacturer′s instruction (Miltenyi Biotech). The resulting cell population was > 95% pure as assessed by flow cytometric analysis using anti CD3 and CD4 antibodies. Intracellular cytokine staining (ICC) The cells from the pleural fluid were directly treated with Brefeldin A (10 mg/mL, Sigma) whereas cultured PFMC were treated four hours before harvesting. Then the cells were fixed with 1% paraformaldehyde and permeabilized with 0.2% saponin (Sigma). Fixed cells were stained with anti-human IFN-γ- FITC and anti-human IL-4 PE at 4°C for 45 minutes, at the concentration recommended by the manufacturer (BD Biosciences). The stained cells were analysed on FACScan (Becton Dickinson) using Cell quest software. Using non-specific isotype matched control assessed the specificity of the staining technique. Cytokine assay Plasma and pleural fluid obtained from the patients were processed within 30 minutes and were stored at -70°C. Plasma, pleural fluid and the supernatants of PBMC and PFMC were thawed at the time of cytokine ELISA. Measurement of IFN-γ and IL-4 was done using the sandwich ELISA kit, according to manufacturer′s instruction (RandD Systems). The average of the duplicate readings was taken as the final concentration and expressed as pg/mL. Statistical analysis Data are presented as the mean ± SEM in both text and in figures. Comparisons between groups were done by Wilcoxon rank sum test. P < 0.05 was considered to be statistically significant. Results In vivo cytokine response To understand the systemic and local in vivo cytokine response, we measured the levels of IFN-γ and IL-4; the prototype cytokines of Th1 and Th2 type immune response respectively in plasma and pleural fluid. The mean IFN-γ and IL-4 levels (± SEM) in plasma (BL) and pleural fluid (PF) of TB and Non-TB patients are shown in [Table - 1]. More than two fold increase in IFN-γ levels in the PF compared with the BL was observed in the TB patients ( P =0.00003). There was no significant increase in the IL-4 levels of PF in TB group. No significant difference was observed between the PF and BL levels of these cytokines in the Non-TB group. In vitro cytokine response To find whether the in vivo pattern of cytokine response is reflected under in vitro conditions, we measured the IFN-γ and IL-4 levels in the supernatants of PBMC and PFMC stimulated with various mycobacterial antigens. The mean levels (± SEM) of IFN-γ and IL-4 when PBMC and PFMC were stimulated with PPD, culture filtrate (CF) and heat killed M. tuberculosis (MTB) are given in [Table - 2]. The IFN-γ levels increased in PFMC when compared with the PBMC in all stimulated conditions. The supernatants of PFMC stimulated with PPD and MTB showed a significant increase in IFN-γ when compared with their control levels (for PPD P =0.03 and for MTB P =0.01). Interestingly, the IL-4 levels were also significantly higher in all the stimulated conditions of PFMC than the control levels. The P values are for PPD P =0.03, for CF P = 0.05 and for MTB P =0.04 [Table - 2]. However, the IL-4 levels in the supernatants of PBMC did not exhibit any prominent variation on stimulation. T-helper cell response To further ascertain the differential nature of cytokine response exhibited by PFMC, we performed both the ex vivo and in vitro intracellular cytokine staining in purified CD4+ T cells for IFN-γ and IL-4, from 10 TB patients. A representative FACS analysis of these data is shown in [Figure - 1]. The ex vivo results showed that the IFN-γ + T cells were significantly higher in pleural fluid than the double positive T cells (IFN-γ+ + IL-4+, P = 0.04) and IL-4 alone positive T cells ( P =0.03). Thus the in vivo response was predominantly of Th1 type [Figure - 1][Figure - 2]. To assess in vitro response, we separated the CD4+ T cells from PFMCs cultured with and without mycobacterial antigens (PPD, CF, MTB) and stained for intracellular cytokines IFN-γ and IL-4. A representative FACS analysis is shown in [Figure - 1]. We observed an antigen specific increase in IFN-γ+ T cells (PPD- P = 0.04, CF - P =0.05 and MTB- P =0.02). Also, the IL-4+ T cells were higher in all cultured conditions, significant for PPD ( P =0.03) indicating that the response was predominantly of Th2 type [Figure - 1][Figure - 3]. Thus, there was an increased Th2 type response to PPD and a mixed response (Th1+Th2) to other antigens. Discussion The present study reports the differences in the localized versus systemic and in vivo versus in vitro immune responses in tuberculous pleuritis. The higher levels of IFN-γ in pleural fluid of the TB patients have been reported in earlier studies.[13],[14] Our results of higher IFN-γ levels in pleural fluid support these observations. In addition, we also demonstrated the selective concentration of IFN-γ+ CD4+ T cells in the pleural fluid. This observation provides the direct evidence that the predominating, protective, local, in vivo immune response in tuberculous pleuritis is of Th1 type. This is concordant with the previous reports that have demonstrated in vitro , the IFN-γ secreting cells derived from PFMC.[15],[16] Similarly, Manca et al have shown that the migration of Th1 cells from blood to the site of infection is more efficient than Th0 cells.[17] The absence of significant increase in the levels of IL-4 and IL-4 secreting T cells in pleural fluid indicates that this cytokine does not have any predominant role at the site of infection as reported earlier.[13],[17] Thus it is evident from our observation and from other reports that there is Th1 response characterized by the presence of IFN-γ+ CD4+ T cells at the site of infection in patients with resistant immune response to tuberculosis. These cells eventually contribute for the bacillary elimination and containment of the infection accounting for the self-limitedness of the disease and hence playing immunoprotective role in tuberculous pleuritis. There are reports that give the profile of cytokine response of specific Mycobacterium reactive human T-cell clones obtained from patients as well as from healthy subjects.[11],[12],[13] The cytokine pattern obtained from such human T-cell clones cultured in vitro, reflected the production of these cytokines only by those specific clones and did not include the contribution of other cells that took part in the immune response to tuberculosis. These studies discord with each other with respect to the T-cell cytokine secretion profile in tuberculosis due to different culture conditions and T helper cell phenotypic variations. The other immune cells and their secreted products, present in milieu are the important factors that influence the sensitization and activation of the CD4+ T cells. Hence we studied the in vitro response of T-helper cells from blood and pleural fluid cultured in the presence of other immune cells to evaluate the overall T-helper cell response operating in milieu of other immune cells. There was no significant difference in in vitro levels of IFN-γ in the antigen-stimulated culture supernatants of PBMC [Table - 2]. This might be due to the recruitment of the antigen specific cells to the site of infection and hence not accessible in the periphery. Moreover, a depressed Th1 response but not an enhanced Th2 response of PBMC to M. tuberculosis antigens is well known in active tuberculosis.[18] In support of this, we observed a decreased Th1 response in PBMC of TB patients when compared with the healthy PPD positive controls (data not shown). In the present study, we observed differences in actual cytokine levels and the ICC positive CD4+T cells when they were cultured in vitro in the milieu of the other cells, isolated from the site of infection. While the established in vivo immune response, as reflected in the cytokine levels of supernatants, was dominated by Th1 type, the ongoing in vitro immune response is predominated by Th2 type, as demonstrated by the preferential increase in IL-4+T cells. Though the quantitative levels of IFN-γ were higher than IL-4 levels in the supernatants, the number of CD4+Tcells secreting IFN-γ was very less compared to IL4 secreting T cells. The contribution of the other immune cells for the secretion of the IFN-γ was less likely as they form only a minor percentage of the PFMC (5% of NK cells and 15% of Tc cells as compared to 72% of CD4+T cells, observed in the present study). Apoptosis of the Th1 cells might be a cause for this observation. Earlier, it has been shown that the activated T-cells are more predisposed to apoptosis.[19] Hence, we speculate that T cells undergoing apoptosis were of Th1 type as the rapid death of Th1 cells by Fas/FasL mediated apoptosis is well-documented.[20] Our study also shows that there is a polarization of cellular response from Th1 type in vivo to Th2 type in vitro. This finding suggests that there may be a mechanism that suppresses IL-4 secretion and subsequent reduction in Th0/Th2 response at the site of infection. This suppression may be due to the high concentration of IFN-γ or due to conditioning by the antigen presenting cells towards Th1 type, thus offering a protective Th1 type response. Alternatively, Th0 cells may show a preferential secretion of type 1 cytokines at the site of disease due to the conditioning microenvironment, which may be absent in in vitro conditions. The absence of these suppressive mechanisms in vitro would have lead to the increased IL-4 levels that support Th2 type response. This is evident from the previous report that showed that the addition of IL-4 favors the development of PPD-reactive clones with Th0/Th2- like cytokine profile.[12] Thus, there seems to be a bias in the T helper cell response operating in in vivo and in vitro in tuberculous pleuritis. Further studies should focus to determine the mechanisms or the factors that lead to the selection of specific cytokine patterns and the reasons for the alterations in immune-responsiveness to tuberculosis infection. Alterations in the in vivo and in vitro immune responses should be considered seriously particularly when we aim at understanding of the immune response of specific cell types or at evaluating antigenic potency of a vaccine against tuberculosis. Acknowledgement Indian Council of Medical Research and Council of Scientific and Industrial Research are acknowledged for providing Senior Research fellowships to CP and KVJ respectively. References
Copyright 2007 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb07006f1.jpg] [mb07006f2.jpg] [mb07006t1.jpg] [mb07006t2.jpg] [mb07006f3.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}