 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Indian Journal of Medical Microbiology, Vol. 26, No. 1, January-March, 2008, pp. 25-28 Original Article Effect of exposure to hydrogen peroxide on the virulence of Escherichia coli Hegde A, Bhat GK, Mallya S Department of Microbiology, Kasturba Medical College, Mangalore - 575 001, Karnataka Date of Submission: 14-Jun-2006 Code Number: mb08004 Abstract Purpose: To eliminate pathogenic bacteria, the host presents conditions that are stressful for bacteria. Oxidative stress arises when the concentration of pro-oxidants like hydrogen peroxide (H 2 O 2 ) and superoxide anion increases to a level over the basal defence capacity of the cell. In the present study, we studied the effect of oxidative stress on the production of certain virulence factors by Escherichia coli .Methods: E. coli was exposed to oxidative stress by growing in the presence of different concentrations of H 2 O 2 . The effect of oxidative stress on the expression of surface hydrophobicity, adherence, haemolysin production, serum resistance and phagocytosis was studied. Results: Oxidative stress caused a significant decrease in the expression of all the virulence factors of E. coli . Conclusions: Synthesis of virulence factors can be significantly altered by oxidative stress and such changes may affect the pathogenicity of E. coli. Keywords: Escherichia coli, oxidative stress, virulence Stress is defined as any deviation from the optimal growth conditions that result in a reduced growth rate. A cell confronted with an abrupt change in its immediate surrounding environment suffers stress. [1] The causes of stress are limitation of essential nutrients such as iron, suboptimal physical conditions such as acidic pH, oxidative stress such as those caused by increased levels of superoxide anion and H 2 O 2 due to oxidative bursts, osmotic stress caused by changes in external osmolarity, that could result in swelling and bursting of the cell in hypotonic environments or in plasmolysis and dehydration in hypertonic conditions. The virulence factors of Escherichia coli are multiple and unusually complex, affecting pathogenicity in combination with one another. The pathogenic processes that operate in a given infection always involve more than one virulence factor. These factors usually interact in so complicated a manner that the precise mechanisms still remain to be defined, but attempts to do so have been made. [2],[3],[4],[5] The common virulence factors include surface hydrophobicity, colonization factor, capsule, serum resistance, resistance to phagocytosis, haemolysins, enterotoxins and siderophores. In the present study, we determined the effect of oxidative stress on the adherence, surface hydrophobicity, hemolysin production, serum resistance and phagocytosis of E. coli . Materials and Methods Bacterial strains Two standard strains of E. coli Microbial Type Culture Collection (MTCC, India) 723, E. coli MTCC 729 procured from MTCC and two clinical isolates of E. coli , E. coli A 030 (isolated from blood of a septicaemic patient) and E. coli A 105 (isolated from urine of a patient with symptomatic urinary tract infection) were used in the present study. The clinical isolates were characterized. Preparation of the bacterial inoculum Each bacterial strain from nutrient agar slope was subcultured twice in chemically defined medium (CDM) of Snyder and Koch, [3] each time at 37 °C for 24 hours. After the final subculture, the cells were washed three times in sterile physiological saline (0.85% w/v NaCl) and finally suspended in saline to get a solution of OD 600 0.1 (corresponds to approximately 1 x 10 8 cells/mL, confirmed by surface plating on nutrient agar). Effect of oxidative stress CDM containing three different molar concentrations of H 2 O 2 (0.1, 0.2 and 0.3 mm) was prepared. [4] These media were dispensed aseptically in 10 mL quantities in three 100 mL Erlenmeyer flasks. Bacterial suspension prepared as described above was used for inoculating the culture media, to achieve an initial concentration of approximately 1 x 10 6 cells/mL in each flask. The flasks were incubated at 37 °C in a rotary water bath at a speed of 160 rpm for 24 hours. The cultures were centrifuged at 3000xg for 10 minutes. The supernatant was discarded and the packed cells were used for the determination of surface hydrophobicity, adherence, haemolysin production, serum resistance and phagocytosis. Effect of stress on the surface hydrophobicity of E. coli The effect of oxidative stress on the surface hydrophobicity of E. coli was studied by quantitative surface hydrophobicity assay, [6] Samples of cultures prepared as described above were washed with sterile phosphate buffered saline (PBS) and then suspended in PBS to get a density of 0.3 at OD 600 (OD Initial). Three millilitres of bacterial suspension was mixed with 0.3 mL of p-xylene and vortexed for 1 minute and left for 30 minutes at room temperature. The hydrophobicity index was calculated by applying the following formula:
Effect of stress on bacterial adherence to epithelial cells The effect of oxidative stress on adherence properties of E. coli was studied by bacterial adherence assay. [7] Bacterial inoculum was prepared as described above. One millilitre of bacterial culture containing about 1 x 10 6 cells/mL was mixed with 1 mL of vaginal epithelial cell suspension containing about 1 x 10 5 cells/mL and incubated at 37 °C for 30 min in a shaker water bath. The cells were centrifuged and the supernatant was filtered to remove free bacteria. The cells were air dried, alcohol fixed and stained for 2 minute with dilute carbol fuchsin. The cell preparation was then examined with oil immersion objective. The number of bacteria adherent to 50 cells were counted and mean was calculated. Effect of stress on the production of haemolysin Effect of oxidative stress on the production of haemolysin by E. coli was studied by the quantitative a-haemolysin assay. [8] Samples of cultures prepared as described above were centrifuged at 3000xg for 10 minutes. The supernatant was diluted in calcium saline and 1 mL of it was mixed with 1% (v/v) sheep erythrocyte suspension and incubated at 37 °C for 1 hour. At the end of the incubation period, 2 mL of saline was added to each tube exhibiting partial haemolysis. The tubes were centrifuged at 1500 rpm for 10 minutes to pellet the unlysed erythrocytes. The supernatant fluid was separated and the OD 540 was determined using the control tube as blank. The 50% haemolysis standard was prepared by mixing 1 mL of 1 % (v/v) sheep erythrocyte suspension and 3 mL of diluent without sodium chloride. Inverse of the dilution which caused 50% lysis was recorded as HU 50 (50% haemolytic units). Effect of stress on serum resistance of E. coli Effect of oxidative stress on the serum resistance of bacteria was evaluated by quantitative serum bactericidal assay. [9] Serum was obtained from healthy donors on the day of each test. Bacterial suspension (0.5 mL) was mixed with 1.5 mL of fresh undiluted serum and incubated at 37 °C. The viable count was determined at the beginning of incubation (0 h) and after 1, 2, 3 h of incubation by surface plating. Each bacterium was tested at least three times. The number of bacteria killed was calculated by applying the following formula:
Effect of stress on the phagocytosis of E. coli The effect of oxidative stress on the phagocytosis of E. coli was studied using polymorphonuclear leukocytes (PMNL). [10] One millilitre of mixture containing 425 μL of E. coli (1 x 10 7 cells/mL), 425 μL PMNLs (5 x 10 6 cells/mL) and 150 μL of healthy human serum (final concentration of 15%v/v) was made in polystyrene tube and rotated slowly at 37 °C for 1 hour. From this mixture, 0.1 mL was added to 9.9 mL of distilled water to lyse the PMNLs and the viable count was determined by surface plating on nutrient agar. A tube of E. coli without PMNLs treated similarly was used as control. The plates were incubated at 37 °C for 48 hour and the extent of killing was determined by using the following formula:
Statistical analysis The data were analysed using the statistical software ′SPSS 11.0′. The means and standard deviation of each of the parameter were obtained. The Kruskal-Wallis anova was used to compare between groups. A statistical significance of the comparison between groups was followed by a multiple comparison criterion to identify the group that was significantly different from the rest. Results Oxidative stress had a significant effect on the surface hydrophobicity of E. coli [Table - 1]. All the strains of E. coli showed a significant decrease in surface hydrophobicity when subjected to oxidative stress by exposure to hydrogen peroxide (H 2 O 2 ). There was a 30 to 50% decrease in the cell surface hydrophobicity of all the three strains of E. coli when exposed to 0.2 mm of H 2 O 2 . Escherichia coli exposed to 0.3 mm H 2 O 2 showed 70% decrease in cell surface hydrophobicity. Oxidative stress had an inhibitory effect on the adherence of E. coli (see [Figure - 1]). E. coli strains showed significant decrease ( P < 0.001) in adherence on exposure to oxidative stress. A significant decrease ( P < 0.001) in adherence was observed with exposure to increasing concentrations of H 2 O 2 in all the four strains of E. coli [Figure - 1]. Oxidative stress had a profound effect on the production of haemolysin by all the strains of E. coli [Table - 2]. Exposure to H 2 O 2 (0.02 and 0.03 mm) caused a significant decrease in the amount of haemolysin produced by all the strains of E. coli . Oxidative stress caused a steep reduction in the amount of haemolysin synthesized by E. coli strain A030. No hemolysin was detected in three strains of E. coli exposed to 0.3 mm H 2 O 2 . Oxidative stress caused a decrease in serum resistance of both the E. coli strains [Table - 3]. The decrease in serum resistance was significant in E. coli MTCC 729 on exposure to 0.02 and 0.03 mm H 2 O 2 . Exposure to 0.1 mm H 2 O 2 did not cause a significant change in serum resistance of this strain. On the other hand, the uropathogenic E. coli A105 showed a significant decrease ( P < 0.05) in serum resistance when exposed to 0.1, 0.2 and 0.3 mm H 2 O 2 . It is clear from the results that exposure to oxidative stress caused a significant decrease in serum resistance of E. coli . Exposure to oxidative stress significantly increased ( P < 0.05) the susceptibility of E. coli to phagocytic killing by PMNLs [Table - 4]. Escherichia coli strain MTCC 729 exposed to 0.2 mm H 2 O 2 and 0.3 mm H 2 O 2 was significantly more ( P < 0.05) susceptible to phagocytic killing. Discussion During infection, E. coli is subjected to different kinds of stress. The results of the present study clearly indicate that oxidative stress may result in changes that may influence the virulence of E. coli . In the present study, we observed that oxidative stress by exposure to H 2 O 2 had an inhibitory effect on the cell surface hydrophobicity and adherence of E. coli. A recent study showed that adherence and surface hydrophobicity of E. coli decreases when grown in Mueller Hinton broth containing oxidizing agents, melatonin and vitamin E. [11] The above finding supports the view that oxidative stress plays an important role in modifying the surface characteristics of E. coli , which could affect the micro-organism′s capacity to adhere to epithelia. Surface hydrophobicity of micro-organisms is one of the virulence factors which contributes to the adherence of micro-organisms to host tissue. [12],[13] Previous studies have shown a correlation between surface hydrophobicity and in vitro adherence to epithelial cells. [14],[15],[16] We observed that any change in the surface hydrophobicity of bacteria resulted in a corresponding change in bacterial adherence to epithelial cells. Oxidative stress also caused a decrease in haemolysin synthesis in both the strains of E. coli . This again could be due to repression of hly genes by H 2 O 2 . Oxidative stress caused an increase in serum susceptibility of E. coli . The present study showed that oxidative stress caused by exposure to H 2 O 2 increased susceptibility of E. coli to phagocytosis. A previous study showed that catalase activity can be induced in several organisms on exposure to exogenous H 2 O 2 . [17] Catalase is thought to be an antioxidant enzyme which protects the bacteria during oxidative stress. It has been reported that gonococci pretreated with H 2 O 2 are significantly more resistant to neutrophils than control bacteria. [17] Such resistance seems most likely to be due to increased formation of catalase. However, we observed increased susceptibility of E. coli to phagocytosis on exposure to H 2 O 2 . The difference in the result could be due to the induction of oxidative stress. Earlier workers have used more than 0.5 mm of H 2 O 2 . [18] It could be that the concentrations of H 2 O 2 used in this study may not have been enough to induce sufficient catalase. Phagocytosis of micro-organisms is a major defence mechanism in humans. Resistance to phagocytosis is an important virulence mechanism of E. coli strains isolated from cases of intestinal and extra-intestinal diseases. Phagocytosis and intracellular killing of E. coli are crucially determined by the surface properties of the bacterial cell. [18] In the present study, we observed that oxidative stress in E. coli could significantly influence the virulence factors of the bacterium. This, in turn, may influence the pathogenesis of E. coli infections. References
Copyright 2008 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb08005f1.jpg] [mb08005t2.jpg] [mb08005t1.jpg] [mb08005t3.jpg] [mb08005t4.jpg] |
| |||||||||
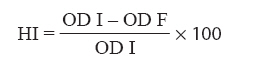
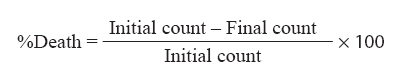

![[Table - 1]](/showimage?mb/photo/mb08005t1.jpg){kind=link}
![[Figure - 1]](/showimage?mb/photo/mb08005f1.jpg){kind=link}
![[Table - 2]](/showimage?mb/photo/mb08005t2.jpg){kind=link}
![[Table - 3]](/showimage?mb/photo/mb08005t3.jpg){kind=link}
![[Table - 4]](/showimage?mb/photo/mb08005t4.jpg){kind=link}