 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
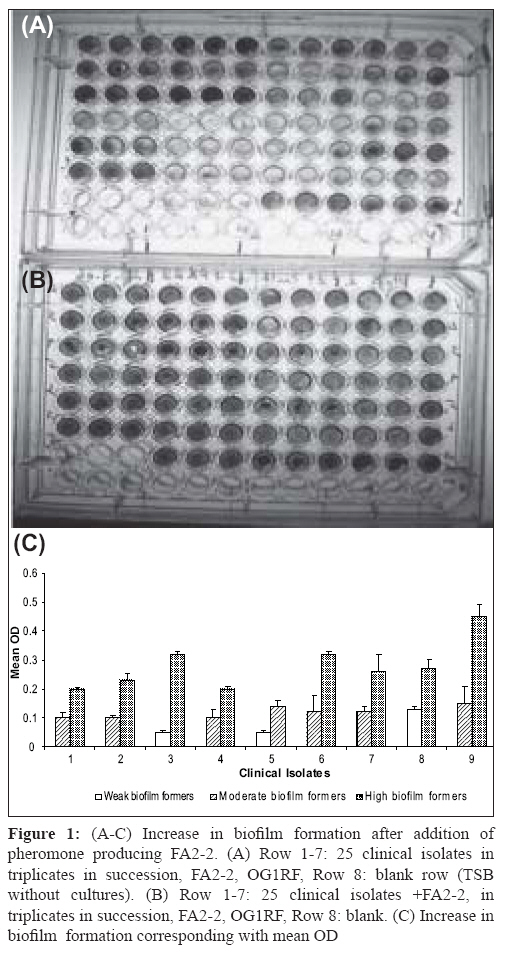
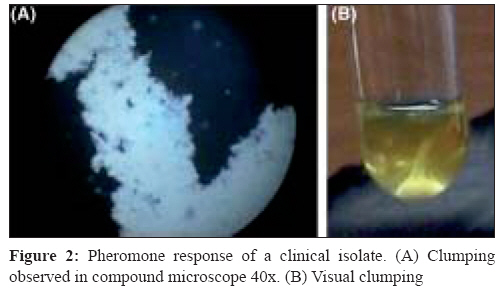
Indian Journal of Medical Microbiology, Vol. 26, No. 3, July-September, 2008, pp. 248-251 Brief Communication Assessment of pheromone response in biofilm forming clinical isolates of high level gentamicin resistant Enterococcus faecalis Jayanthi S, Ananthasubramanian M, Appalaraju B Department of Biotechnology, PSG College of Technology, Coimbatore - 641 004, Tamil Nadu Date of Submission: 09-Jul-2007 Code Number: mb08077 Abstract Twenty five clinical isolates of high level gentamicin resistant Enterococcus faecalis were tested for their biofilm formation and pheromone responsiveness. The biofilm assay was carried out using microtiter plate method. Two isolates out of the 25 (8%) were high biofilm formers and 19 (76%) and four (16%) isolates were moderate and weak biofilm formers respectively. All the isolates responded to pheromones of E. faecalis FA2-2 strain. On addition of pheromone producing E. faecalis FA2-2 strain to these isolates, seven of 19 (37%) moderate biofilm formers developed into high biofilm formers. Similarly one of the 4 (25%) weak biofilm formers developed into high level biofilm former. Twelve (48%) of the 25 isolates were transconjugated by cross streak method using gentamicin as selective marker. This proves that the genetic factor for gentamicin resistance is present in the pheromone responsive plasmid. Among these twelve transaconjugants, seven isolates and one isolate were high biofilm formers on addition of E. faecalis FA2-2 and prior to its addition respectively. Out of the total 25 isolates, eight transconjugants for gentamicin resistance could turn to high biofilm formers on addition of the pheromone producing strain. All the isolates were resistant to more than two antibiotics tested. All the isolates were sensitive to vancomycin. The results indicate the significance of this nosocomial pathogen in biofilm formation and the role of pheromone responding clinical isolates of E. faecalis in spread of multidrug resistance genes. Keywords: Biofilm, enterococcus faecalis, pheromone Enterococci are gram positive bacteria that normally inhabit the gastrointestinal tract of many animals including humans. [1] However, when they colonise habitats where they are not normally found, these opportunistic bacteria can become pathogens. Enterococcal infection incidence, especially hospital acquired, has dramatically increased over the last 20 years. Enterococci have been reported recently as a major cause of nosocomial infections [2] being increasingly detected in blood-stream and urinary tract infections and in infected surgical sites. Enterococcus faecalis is responsible for approximately 80-90% of all enterococcal infections. The intrinsic antibiotic resistance of enterococci, coupled with their promiscuity in acquisition and dissemination of genetically mobile antibiotic resistance elements, presents serious challenges to the treatment of enterococcal infections. Infection-derived isolates of E. faecalis have been shown to form biofilms in vitro . [2] Furthermore, E. faecalis is often isolated from biofilms on the surfaces of various indwelling medical devices associated with chronic infection. [3],[4] Resolution of these infections is complicated by antibiotic resistance that can be associated with pheromone responsive plasmids. Transfer of some enterococcal conjugative plasmids from donor to recipient cells is induced by signaling peptides (pheromones). The present study was carried out to assess the potential of the clinical isolates of E. faecalis to respond to the pheromones and also to the extent these isolates can form biofilm. Materials and Methods Samples Biofilm assay Clumping assay Pheromone inducing biofilm assay Conjugation method Results A total of 25 clinical isolates of high level gentamicin resistant Enterococcus faecalis were isolated from different clinical samples [Table - 1]. All these isolates were resistant to more than two antibiotics tested (data not shown). All the isolates were sensitive to vancomycin. On screening the isolates for biofilm, two isolates out of 25 isolates (8%) showed high biofilm formation. Nineteen (76%) and four (16%) isolates were moderate and weak biofilm formers respectively. The categorisation of biofilm was done based on the OD value of E. faecalis standard strain OG1RF. The OG1RF strain showed the mean OD value of 0.12 at 490nm. Those in and above this value were categorised as moderate biofilm formers. Those below this value were categorised into low or non biofilm former [Table - 2]. Also those above 0.2 OD value were considered as high biofilm formers. No significant correlation between biofilm forming nature and source of the clinical isolates was observed. All the twenty five isolates tested for pheromone response to FA2-2 strains showed clumping [Figure 1A]. Clumping was both observed visually by naked eye and microscopically. On addition of pheromone producing culture E. faecalis FA2-2 strain [Figure 1B], 10 (40%) out of 25 isolates were found to be high biofilm formers. One of four (25%) weak biofilm formers developed into high level biofilms. Similarly seven of 19 (35%) moderate biofilm formers developed into high biofilm formers [Figure 1C]. No significant change was observed in the OD values of the already two high biofilm formers and remaining 12 moderate biofilm formers. However, all the three remaining weak/non biofilm formers turned to moderate biofilm formers Twelve out of the 25 isolates were tested by cross streak mating developed transconjugants that support the transfer of high level gentamicin resistance to the donor strains. Among these, seven isolates and one isolate were high biofilm formers on addition of E. faecalis FA2-2 and prior to its addition respectively. Apart from them one transconjugant turned from weak to moderate on addition of FA2-2 [Figure - 2]. The remaining four isolates showed no change in biofilm formation Discussion Enterococcus faecalis is one of the major etiologies of nosocomial infections. Gentamicin is being used along with a beta lactam antibiotic. High level gentamicin resistance poses a problem in treating the patients which leads to use of vancomycin as the last resort. The potential relationship between biofilm formation and clinical implications of E. faecalis isolates has been established earlier. [8] Tissue culture plates made of polystyrene were found to be most sensitive, accurate and reproducible screening method for detection of biofilm formation in clinical isolates. [9] In this study, 96 well polystyrene microtiter plates were used. The detection of two high biofilm (8%) formers shows a threat for nosocomial infection. One of the isolates was from blood and other from pus. Here we have used E. faecalis OG1RF strain as a standard for biofilm formation. It has been reported to be a moderate biofilm former. [10] The detection of clumping response in all the clinical isolates underlines the presence of pheromone responding plasmids that carry the virulence factors. Each type of plasmid responds to specific pheromone. E. faecalis FA2-2 is without any plasmid and is expected to secrete different types of pheromones. Hence, it is likely that these isolates harbour different types of plasmids. Most pheromone-responsive plasmids found in Enterococcus faecalis exhibit a narrow host range and transfer between E. faecalis strains at a high frequency (10 0 to 10 -2 per donor cell) within a few hours during broth mating. [11] The plasmids confer a mating response to a small peptide (i.e., a sex pheromone) secreted by potential recipient cells. This mating signal induces the synthesis of a surface aggregation substance that facilitates the formation of mating aggregates. Plasmid-free recipients secrete multiple sex pheromones, each specific for a donor harbouring a related pheromone-responsive plasmid. Once a plasmid is acquired by the recipient, secretion of the related pheromone ceases, whereas other unrelated pheromones continue to be produced. Determinants encoded on pheromone-responsive plasmids include those for haemolysin, bacteriocin, and resistance to UV light and antibiotics. [12] Interestingly, though clumping occurred in all these biofilm formers, not all developed into high level biofilm formation. The seven moderate biofilm formers almost doubled their OD value on addition of pheromone. Whereas the remaining twelve moderate biofilm formers showed no significant change in their OD values. Addition of pheromone to biofilm formers has yielded interesting results. Some of the isolates of moderate and weak/non biofilm formers turned to high biofilm formers. This supports the earlier views on pheromone induced biofilm formation. [13] The other isolates that showed no increase in biofilm formation for the addition of E. faecalis FA 2-2 could be due to the lower concentration of the specific pheromone to the particular isolate in pheromone of FA2-2. [14] The twelve transconjugants developed also highlight the efficiency of the clinical isolates to disseminate the resistance factor. This proves the genetic factor for gentamicin resistance present in the pheromone responsive plasmid. For the remaining non conjugant isolates, the use of gentamicin as selective marker could probably have missed the transconjugant for other virulence factors. Eight out of the 12 transconjugants have shown an increase in their biofilm forming ability when cultured in the presence of the pheromone producing FA 2-2. Conjugative plasmid transfers in the biofilm forming isolates have been reported in E. faecalis . [15] The present study highlights the enhancement of biofilm formation on pheromone addition to the isolates. Of the nineteen moderate biofilm formers, seven of them turned to high biofilm formers and six of them were capable of conjugation towards transferring gentamicin resistance. Out of the total 25 isolates, eight transconjugants for gentamicin resistance could turn to high biofilm formers on addition of pheromone producing strain and hence are a potential risk in terms of spreading the virulence factors and in developing drug resistance. The enhanced antibiotic tolerance resulting from growth in a biofilm state, coupled with the intrinsic antibiotic resistance of the organism as well as its propensity to serve as a reservoir for the dissemination of antibiotic resistance genes, suggests that biofilm formation of E. faecalis in a hospital setting may be particularly problematic. Acknowledgement The authors thank University Grants Commission, New Delhi for funding the project.References
Copyright 2008 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb08077f1.jpg] [mb08077t2.jpg] [mb08077f1a&b&c.jpg] [mb08077f2.jpg] [mb08077t1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}