 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
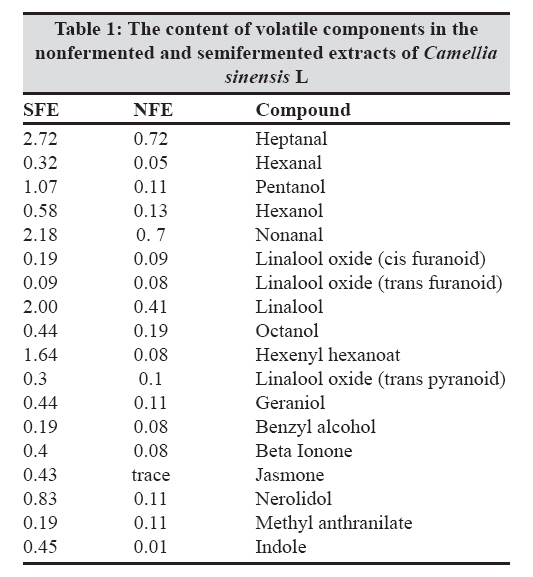
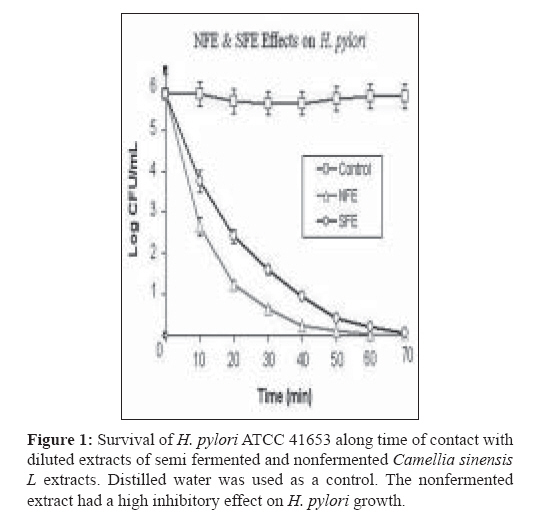
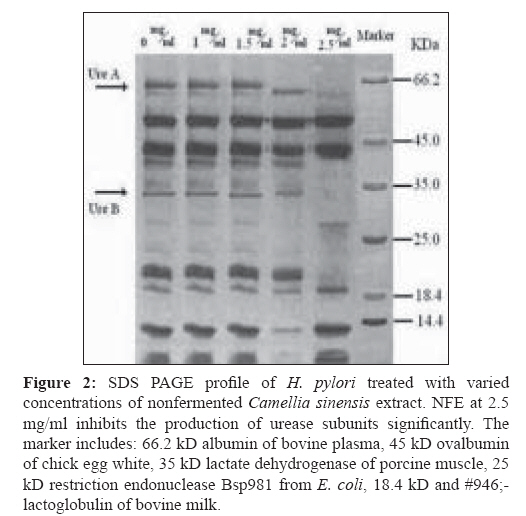
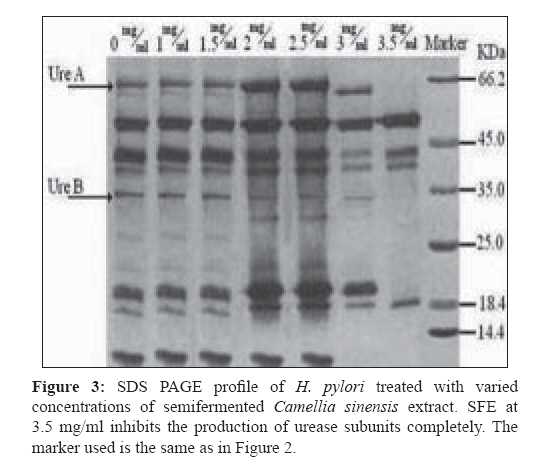
Indian Journal of Medical Microbiology, Vol. 27, No. 1, January-March, 2009, pp. 30-34 Original Article In vitro inhibition of Helicobacter pylori urease with non and semi fermented Camellia sinensis Shoae Hassani AR, Ordouzadeh N, Ghaemi A, Amirmozafari N, Hamdi K, Nazari R Faculty of Medicine, Golestan University of Medical Sciences, Gorgan Date of Submission: 01-May-2008 Code Number:mb09007 Abstract Purpose: Helicobacter pylori is the etiological agent in duodenal and peptic ulcers. The growing problem of antibiotic resistance by the organism demands the search for novel compounds, especially from natural sources. This study was conducted to evaluate the effect of Camellia sinensis extracts on the urease enzyme that is a major colonization factor for H. pylori.Methods: Minimum inhibitory concentrations of nonfermented and semifermented C. sinensis methanol: water extracts were assessed by broth dilution method. Examination of the urease function was performed by Mc Laren method, and urease production was detected on 12% SDS polyacrylamide gel electrophoresis from whole cell and membrane bound proteins. Results: Both extracts had inhibitory effects against H. pylori and urease production. At a concentration of 2.5 mg/ml of nonfermented extract and 3.5 mg/ml of semifermented extract the production of Ure A and Ure B subunits of the urease enzyme were inhibited completely. A concentration of 4 mg/ml of nonfermented and 5.5 mg/ml of semifermented extract were bactericidal for H. pylori. Conclusions: C. sinensis extracts, especially the nonfermented, could reduce H. pylori population and inhibit urease production at lower concentrations. The superior effect of nonfermented extract is due to its rich polyphenolic compounds and catechin contents. Keywords: Camellia sinensis, catechin, Helicobacter pylori, urease Young shoots of Camellia sinensis plants are the source of tea, the commonest beverage in the world. Immediately after harvest, the tea leaves are heated with rolling to inactivate the enzyme polyphenol oxidase, capable of oxidizing the catechins to oligomeric and polymeric derivatives such as theaflavins and thearubigins. [1] Green or nonfermented tea is thus less fermented and has the highest quantity of catechins that are chemically defined as flavan 3 ols. When the enzyme is allowed to act longer, the tea will ferment more and most of the catechins will be converted to theaflavins and thearubigins that give the aroma and color to semifermented tea. [2] The phenolic content of nonfermented C. sinensis is between 50-100 mg polyphenols per cup, depending on the species, harvesting variables, and brewing methods. [3] If green tea is directly heated, the catechins are destroyed and lacks polyphenols. In Asian cultures that consume a lot of green tea it is brewed under the steam of boiling water, thus, retaining the catechins. [2] Until recently, there was a paucity of evidence of the antimicrobial activity of C. sinensis . Although, in the recent past there are several reports of its antibacterial effects in vitro and in vivo, the latest series of well-conducted systematic studies, mostly from Japan, suggest of C. sinensis extracts have several useful antimicrobial effects. [4] Helicobacter pylori that infects various areas of the stomach and duodenum is a major etiological agent of peptic ulcers, duodenitis, and perhaps some malignancies. [5] The occurrence of strains resistant to clarithromycin is a cause of concern, especially in geographical areas with a high incidence H. pylor infection. [6] Therefore, it is important to search for novel therapeutic agents, other than antibiotics, that are both highly effective and safe. The only known niche of H. pylori is the gastric mucosa, where large pH fluctuation occurs. H. pylori enters the stomach, and with the help of the urease enzyme migrates from the acidic lumen into the gastric mucus layer, where the pH is thought to vary between 4 and 6.5 and where occasional acid shocks of pH as low as 2 can occur. [7] More than 50% of the human population in the world and more than 80% of people in developing countries are infected with H. pylori, increasing the risk or oral transmission. [ 8 ] Urease is an extracellular cell bound enzyme of H. pylori constituting 5-10% of the total cell protein with a molecular weight of approximately 580 kD comprising of six 66 kD (Ure A) and six 32 kD (Ure B) subunits. [9] This study was conducted to evaluate the anti H. pylori effect and urease inhibition by semifermented and nonfermented Camellia sinensis extracts. Materials and Methods Extraction of non and semifermented Camellia sinensis Young shoots of Camellia sinensis L were harvested from the hilly regions of Lahijan city in the south coast of Caspian Sea in Iran. Semifermented and nonfermented teas were produced from the harvested shoots by the rolling and heating process that include a fermentation period between rolling and heating for the semifermented tea. The semi and nonfermented C. sinensis L were stored in plastic bags and transported to the microbiology laboratory of Sciences and Research campus of IAU situated in Tehran. Semi and nonfermented C. sinensis L were powdered in a blender and then were extracted with methanol: water mixture (62.5:37.5 v/v) in the clevenger extractor as follows: dry powders (500 mg) were weighed in test tubes. A total of 10 mL of 62.5% aqueous methanol was added to the tubes and the suspensions were slightly stirred. Tubes were sonicated for 5 minutes and centrifuged for at 1500 g for10 mins. The supernatants were collected and the powders were reprocessed twice in the clevenger extractor. The nonfermented (NFE) and semifermented (SFE) extracts were concentrated by evaporation to one-fifth the volume, filter sterilized with 0.21µ paper, and stored at 4 °C. Gas chromatography of non and semifermented Camellia sinensis GC analyses were performed using a Shimadzu 17A gas chromatograph (Shimadzu, Japan) equipped with a flame ionization detector and a 60 m x 0.25 mm (I.D.) DB-WAX (J and W Scientific, Folsom, CA) fused-silica capillary column. The operating conditions were as follows: oven temperature, 40 °C, 3 °C /min rise to 220 °C; carrier gas, helium; and flow rate, 1.0 ml/min. The retention times and peak areas of the eluted volatile components were integrated with a chromatopack integrator. Gas chromatographic-mass spectrographic analysis GC-MS analysis was carried out on a Hitachi M-808 double-focusing instrument equipped with a Hewlett-Packard 5890 gas chromatograph. The GC conditions were identical to those of the above analytical GC runs. Mass spectral data were acquired and processed by a built-in computer system (M-0101) developed by Takasago International. The components of the distillate were identified by comparing their GC retention times and MS fragmentations with those of the authentic samples. Bacterial culture H. pylori ATCC 41653 purchased from Persian Type Culture Collection in Tehran was cultured on brucella broth, Oxoid and Columbia agar, Oxoid (Basinstoke, UK) at 37 0 C with 7% defibrinated sheep blood incubated in 10% CO 2 . For antimicrobial susceptibility test, Mueller Hinton broth (Difco, France) was used in broth dilution test. Urease activity was detected in urea broth (Merck, Germany). For the inhibition of other microorganisms and prevention of contamination, 10 μg of vancomycin and 5 μg amphotericin B (Sigma Chemicals, France) were added to the media.[10] Antimicrobial susceptibility test Broth dilution method was used to evaluate the minimum inhibitory concentrations (MICs) and minimum bactericidal concentrations (MBCs) of NFE and SFE. Various concentrations of the extracts were added to 10 ml brucella broth containing 7% defibrinated sheep blood, inoculated with 10 6 colony forming unit (CFU)/mL of H. pylori suspensions and incubated in 10% CO 2 incubator. After 5 minutes at room temperature, surviving CFUs were counted on Columbia agar incubated in 10% CO 2 after 48 hours. Disk diffusion method was used to determine the zone of inhibition. The 72 hour H. pylori culture from brucella broth incubated in 10% CO 2 was centrifuged for 10 minutes at 3000 g. The pellet was suspended in 5 ml of peptone water to achieve a concentration of 10 6 CFU (with Shimadzo UV 120-1 spectrophotometer). Hundred microliters of this suspension was inoculated onto Mueller Hinton Agar (MHA) with a cotton swab, and then 20-60μg of NFE and SFE (100 mg/ml) were added to 6 mm paper disks. The disks were dried slowly in darkness and a disk of each extract was placed on MHA supplemented with 7% defibrinated sheep blood and incubated in the 10% CO 2 incubator for 48 hours at 37 0 C. Metronidazole disk (8 µg) was used as a positive control and distilled water disk as a negative control. Each experiment was duplicated along with controls. Urease activity assay Mc Laren method was used to assess urease function. [11] Some colonies of H. pylori from 72-hour cultures on Columbia agar with 7% defibrinated sheep blood were picked and suspended in peptone water, to achieve a concentration of 10 6 CFU/mL. Hundred microliteres of this suspension was inoculated into 24 tubes of brucella broth with 7% defibrinated sheep blood. These cultures were treated with 0-6 mg/ml of NFE and SFE and were incubated at 37 0 C for 48 hours. The viable bacteria from the last concentration were centrifuged for 10 minutes at 3000 g. The harvested cells were added to urea broth and vortexed for 10 minutes. A change in the color of urea broth signified the presence of active urease enzyme. If there was no observable color change in the urea broth, this process was repeated, and if there was still no color development, then, the urease activity would be reported as negative. Urease production assay on SDS PAGE For SDS PAGE analysis, H. pylori treated cells with different concentrations of NFE and SFE were centrifuged for 5 minutes at 5000 g and the pellets were resuspended in 30 mM Tris HCl, pH 8.1. The suspensions were recentrifuged for 10 minutes at 10000 g and the pellet vortexed in 200 µl of 20% sucrose in Tris HCl, after which, these cells were resuspended in phosphate buffer at pH 7. The cells were incubated on ice with 33 mg/L lysozyme for 30 minutes and disrupted by sonication for 20 seconds. Soluble and insoluble fractions were separated by centrifuging for 15 minutes at 15,000 g. Hundred microliters of the supernatant was mixed in 3LUG sample buffer and a SDS PAGE analysis performed according to the method of Laemmli on 12% polyacrylamide gel. [12] Gels were stained with Coomassie brilliant blue for quantification. Statistical analysis All experiments were carried out in triplicate, and each sample was analyzed in duplicate. The results were expressed as mean ± S.D (standard deviation) and analyzed statistically by ANOVA test ( P < 0.01). Results Extracts of nonfermented and semifermented C. sinensis L from Lahijan city in Iran, were compared by means of GC [Table - 1]. The amounts of almost all the volatile components including alcoholic, aromatic constituents hydrolyzed by the enzyme increased as a result of semifermentation of catechins. The content of each component is expressed by the ratio of each peak area of the internal standard of the gas chromatogram On comparing NFE and SFE, the growth inhibitory concentration of NFE on H. pylori was lower indicating that its antimicrobial activity was better than SFE. The diameter of the inhibitory zone at a concentration of 4 mg/ml was 22.5 mm for NFE and 18 mm for SFE. A remarkable finding when compared with the positive control (Metronidazole, 8 µg) that had a 21 mm inhibitory zone. The bactericidal concentrations of NFE and SFE on H. pylori were 4 and 5.5 mg/ml, respectively. The bactericidal effect of both the extracts was obvious and NFE was faster in inhibiting the growth of H. pylori . The time taken to achieve a viable cell count below 1 log of CFU/mL at 5 mg/ml was 30 minutes with NFE and 40 minutes with SFE [Figure - 1]. Growth of H. pylori was observed at a concentration of 4 mg/ml of SFE and 3 mg/ml of NFE. A significant decrease in bacterial number was observed at these concentrations of the extracts in contrast to untreated cultures [Figure - 1]. In the Mc Laren test, inhibition of urease activity was seen at a concentration of 1.5 mg/ml of NFE and at 2.5 mg/ml of SFE. The analysis of SDS PAGE revealed that, at 2.5 mg/ml of NFE, both, the 65 kD band of the larger subunit of urease (Ure A) and the 32 kD band of its smaller subunit (Ure B) completely disappeared. At 2 mg/ml, these two bands did appear but were smaller than those at previous concentrations [Figure - 2]. At 3 mg/ml of SFE, the urease enzyme subunits were clearly observed but produced in lower quantity. Urease production was inhibited completely by 3.5 mg/ml of SFE [Figure - 3]. Discussion Since urease of H. pylori is an essential enzyme for its colonization, we focused on natural sources that could inhibit the activity of this enzyme. Tea consumption is known to be not only beneficial in providing immunity against many diseases but also in H. pylori infection as authenticated by our results. To determine MICs of the non and semifermented C. sinensis L extracts, we used broth dilution test that was a simple, rapid, and cost-effective. Based on the National Institute of Health and evaluation of laboratory standards in Wales (NHS), our method was modified to the needs of H. pylori. [10] C. sinensis L is rich in catechins like epicatechin, epigallocatechin, epicatechin gallate, and epigallocatechin gallate. Nonfermented C. sinensis contains a high level of the catechins though these molecules are lost during semi fermentation processes. In comparison between NFE and SFE , the former had higher antimicrobial activity against H. pylori despite the more volatile components that exist in SFE as a result of semi fermentation process [Table - 1]. Studies conducted over the last 20 years have shown that the polyphenolic catechins, particularly epigallocatechin gallate and epicatechin gallate, can inhibit the growth of a wide range of Gram-positive and Gram-negative bacterial species with moderate potency. The results of the Mc Laren test showed that 1.5 mg/ml of NFE and 2.5 mg/ml of SFE could inhibit the urease activity of this bacterium completely. Surprisingly, at these concentrations of the extracts the production of urease was constituted and its bands were observed on SDS PAGE [Figure - 2]. These data confirm the direct urease inhibition activity of C. sinensis L extracts in addition to their inhibitory effects on urease production in viable H. pylori cells [Figure - 2] and [Figure - 3]. In 2003, Matsubara et al, found that, tea catechins, particularly epigallocatechin gallate could inactivate urease enzyme. [9] It has been demonstrated that H. pylori lacking urease activity are incapable of causing infection in animal models. Tsujii et al , (1992) confirmed the necessary role of urease in conversion of urea to ammonia and protection of H. pylori from digestion by gastric juice. [13] Eaton et al , (1991) showed that urease has a key role in H. pylori pathogenesis [14] and in 1995 Andrutis et al , explained the relationship between ammonia production in survival and colonization of H. pylori in low acidic pH. [15] In the presense of urease inhibitory or bactericidal concentrations of NFE or SFE in the gastric mucus it is possible to decrease the rate of gastric infection or malignancies caused by H. pylori . It has been reported that C. sinensis catechins have antibacterial activity against foodborne pathogenic bacteria. Also, in 1993 Ikigai et al, had reported that tea catechins could damage the bilayer lipid membrane, therefore catechins could damage H. pylori cell membrane and inhibit its proton pump actions. [16] Damage to the cell membrane can destroy the urease enzyme as its presense is restricted to the cell membrane of H. pylori . Urease production disappeared at 2.5 mg/ml of NFE, while production of urease enzyme was seen at this concentration of SFE on SDS polyacrylamide gel electrophoresis. SFE at 3.5mg/ml had inhibitory effect on urease production of H. pylori [Figure - 2] and [Figure - 3]. Because NFE and SFE can diffuse into the helicobacter cells, they can disrupt the urease of H. pylori cells as soon as it is produced. NFE at 4 mg/ml and SFE at 5.5 mg/ml had bactericidal activity on H. pylori . The reason of this significant difference between the efficacies is the plentiful content of catechin molecules in NFE rather than SFE. Catechins, known for their antioxidant activity, [17] antiinflammatory effect, [18] and inhibition of gastric secretions, [17] contribute in part to this efficacy. Investigations in epidemiological studies have shown an inverse relationship of NFE consumption with risk of gastric malignancies. The risk of stomach cancer decreases with the quantities of tea consumed. [19] Decrease in H. pylori numbers and low urease production inhibit H. pylori colonization and therefore could reduce chronic gastritis, peptic ulceration, mucosa-associated lymphoid tissue lymphoma, and adenocarcinoma of the stomach. In 2000, Kuwahara et al, Shibata et al, and Setiawan et al , concluded that nonfermented C. sinensis could also prevent chronic active gastritis and decrease the risk of stomach cancer. [20] In conclusion, our findings revealed that both semifermented and nonfermented C. sinensis L extracts have anti H. pylori effects. These extracts can inhibit the growth of this bacterium and at lower concentrations inhibit the function and production of the enzyme urease. So, consumption of tea, a safe compound, can reduce the colonization of H. pylori in the gastric mucosa. Acknowledgment The authors wish to thank Miss. Behin Omidi from the laboratory of microbiology in Science and Research branch of IAU for kindly providing the materials for the experiment as well as beneficial information.References
Copyright 2009 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb09007t1.jpg] [mb09007f3.jpg] [mb09007f1.jpg] [mb09007f2.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}