 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
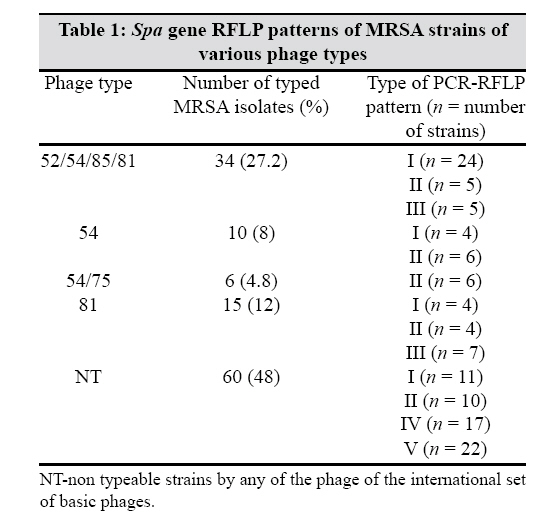
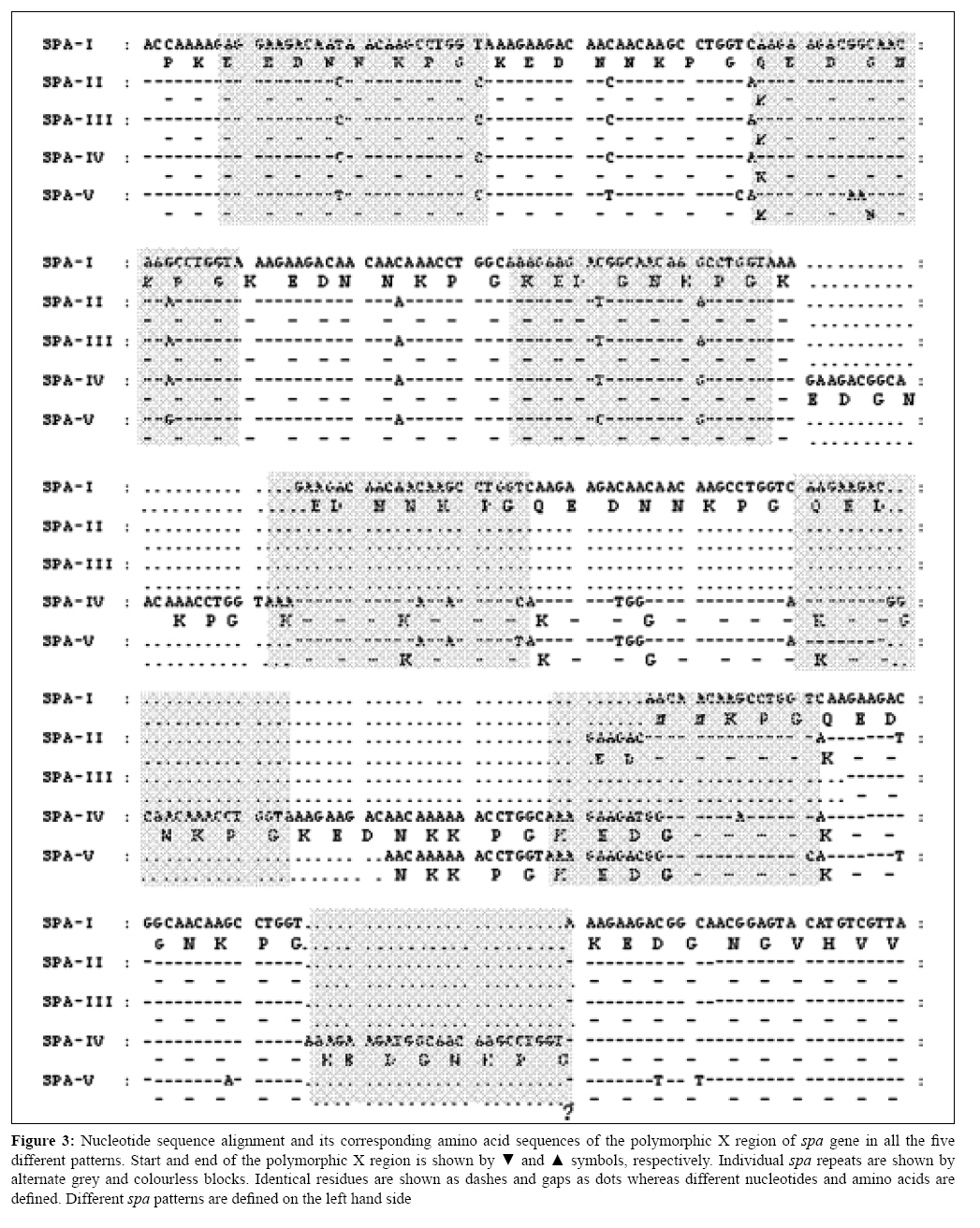
Indian Journal of Medical Microbiology, Vol. 27, No. 2, April-June, 2009, pp. 116-122 Original Article Molecular typing of methicillin-resistant Staphylococcus aureus strains by PCR-RFLP of SPA gene: A reference laboratory perspective Mehndiratta PL, Bhalla P, Ahmed A, Sharma YD Department of Microbiology, National Staphylococcal Phage Typing Center, Maulana Azad Medical College, New Delhi-110 002 Date of Submission: 06-May-2008 Code Number: mb09037 PMID: 19384033 DOI: 10.4103/0255-0857.45363 Abstract Purpose: To characterize methicillin-resistant Staphylococcus aureus (MRSA) strains by molecular typing based on polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) of spa gene and to assess the utility of spa genotyping over bacteriophage typing in the discrimination of the strains.Materials and Methods: Studies were undertaken on 125 MRSA strains representing the most predominant phage types and the non phage typeable strains. Strains were typed by bacteriophage typing and PCR-RFLP of spa gene. DNA sequence analysis of the amplified spa gene fragment of the representative RFLP patterns was performed using standard protocols. Results: All the strains resistant to oxacillin were found to contain mec A gene. Fifty-two per cent of these strains were typeable by the international basic set of 23 phages. Five different PCR-RFLP patterns were observed among 125 MRSA strains. Non phage typeable strains were differentiated into four PCR-RFLP patterns. Sequencing of the spa gene from the representative strains of each RFLP pattern confirmed the length of these restriction fragments due to variation in the 24 bp and the 174 bp tandem repeats. It also revealed the presence of three new spa repeat patterns. Conclusion: The study demonstrates the importance of spa genotyping in the discrimination of MRSA strains, which were otherwise indistinguishable by bacteriophage typing. spa genotyping allowed differentiation of strains within a particular phage type. Nucleotide sequencing of isolates of different PCR-RFLP patterns indicated a correlation between the RFLP patterns of a variable number of tandem repeats and the phage type. The study provides valuable information on the epidemiological characterization of MRSA strains. Keywords: Methicillin resistance, MRSA, PCR-RFLP, spa gene, Staphylococcus aureus Introduction Staphylococcus aureus is one of the most significant pathogen known for causing sporadic infections and epidemics. Methicillin-resistant Staphylococcus aureus (MRSA) strains were first identified in 1961 immediately after the introduction of methicillin in clinical settings. [1] Subsequently, increase in resistance to methicillin among S. aureus isolates has been observed globally. [2],[3],[4] India too has a significant problem with MRSA because there is a steady increase in the occurrence of MRSA strains in its hospitals. [5],[6] This is a cause of concern and requires rapid precise typing of MRSA for epidemiological surveillance and hospital infection control. The bacterial strain typing distinguishes epidemiologically related or clonal isolates from unrelated ones. Epidemiologically related isolates are viewed as descendants from a common precursor and thus will be indistinguishable but recognizably different from unrelated isolates from the same species. Our earlier studies, based on bacteriophage typing, hypothesized that only few related clones are present within various hospitals in India. [6] Bacteriophage typing is subjective and at times the strains are not typeable. This requires alternate strategies to further distinguish these non-typeable MRSA strains. Recently, several investigators have described DNA-based techniques for typing S. aureus strains. Of these, pulse field gel electrophoresis (PFGE) is recognized as the most useful and discriminatory, but it is relatively costly and technically complex. [7] Alternatively, polymerase chain reaction (PCR)-based methods, targeting various genes, have been used successfully for this purpose. [8] Genes coding for two specific proteins, coagulase ( coa ) and protein A ( spa ) have been the most widely used markers for molecular typing as they contain highly polymorphic repeat units. [9],[10] Polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) studies of these genes were found to be quite useful in typing S. aureus strains and have proven absolute typeability, reproducibility and good discriminatory power. [11] The present study was carried out to characterize MRSA strains by molecular typing based on PCR-RFLP of spa gene and to further investigate the utility of spa genotyping over bacteriophage typing in the discrimination of strains.Materials and Methods Collection of MRSA isolates Studies were undertaken on MRSA strains collected for phage typing at the National Staphylococcal Phage Typing Center, Maulana Azad Medical College, New Delhi, India, from various hospitals located in different parts of the country. Bacteriophage typing was carried out using 23 phages from the international set of phages. A total of 125 isolates were selected so as to represent the most predominant phage types and non phage typeable isolates. S. aureus NCTC 6571 and ATCC 43300 were used as negative and positive controls for methicillin resistance, respectively. Detection of methicillin resistance Methicillin resistance was determined for all the S. aureus isolates by the agar screening method [12] using Mueller Hinton agar supplemented with 4% NaCl and 6 µg/mL of oxacillin (Sigma Aldrich, St. Louis, MO, USA). Methicillin resistance was further confirmed by the detection of mec A gene (encoding high resistance to methicillin) by the PCR method, which is described below. [13] Phage typing Phage typing was performed according to the recommendations of Blair and Williams [14] using a basic set of 23 phages at 100x routine test dilution, which were as follows: Group I, 29, 52, 52A, 79, 80; Group II, 3A, 3C, 55, 71; Group III, 6, 42E, 47, 53, 54, 75, 77, 83A, 84, 85; and non-allocated 81, 94, 95, 96. The results were interpreted by using the criteria of differentiating the strains if they showed a difference of two strong reactions. DNA isolation and PCR After an overnight culture on nutrient agar plate, one colony of each sample was suspended in 50μL of water and the suspension was then placed in a 100°C heat block for 15 min. From this suspension, a 5μL volume was directly used as template for PCR amplification of mec A and spa gene fragments. mec A1 (5 ′ - GTA GAA ATG ACT GAA CGT CCG ATA A - 3 ′ ) and mec A2 (5 ′ - CCA ATT CCA CAT TGT TTC GGT CTA A - 3 ′ ) primers were used for the amplification of the 310 bp fragment of the methicillin-resistant gene ( mec A) [13] and primers spa 1 (5 ′ - ATC TGG TGG CGT AAC ACC TG - 3 ′ ) and spa 2 (5 ′ - CGC TGC ACC TAA CGC TAA TG - 3 ′ ) were used to amplify the portion of the spa gene. [11] Amplification was carried out in a Gene Amp PCR system 9700, (Applied Biosystems). The PCR mixture consisted of 1 mM MgCl 2 , 0.2 mM dNTPs (Invitrogen Life Technologies, Carlsbad, CA, USA), 1 unit of taq DNA polymerase (Invitrogen Life Technologies), 1 µM of primers and 5μL of DNA extract in a final volume of 50μL. For spa gene segment amplification, the PCR conditions were as follows: initial denaturation at 94°C for 7 min followed by 35 cycles of denaturation at 94°C for 1 min, annealing at 56°C for 1 min and extension at 72°C for 3 min with a final extension at 72°C for 5 min. The part of the mec A gene was amplified under similar conditions except that the annealing temperature was 60°C and extension was for 40s. The PCR products were subjected to agarose (1.2% for spa fragment and 1.5% for mec A fragment) gel electrophoresis. PCR-RFLP RFLP of the PCR product was carried out by the modified method of Wichelhaus et al. [11] Bands of the PCR product of the spa gene were excised out from the gel and DNA was extracted using the QIAEX-II Gel Extraction kit (Qiagen GmbH, Hilden, Germany) following the manufacturer′s instructions. The DNA was eluted in a 30µL sterile triple-distilled water. Ten microlitres of this was taken and digested with one unit of BSP 143II restriction enzyme (MBI Fermentas GmbH, Germany) at 37°C for 16 hours. Digested fragments were separated on 2.5% agarose gel. DNA sequencing One hundred to 200ng of the above gel-purified PCR product was used for sequencing using an ABI Big Dye Terminator Ready Reaction kit Version 3.1 (PE Applied Biosystems, CA, USA) under the same conditions as described before. [15] Templates were purified and sequenced on an ABI Prism 310 Genetic Analyzer (PE Applied Biosystems). DNA sequence analysis The sequences obtained were analysed and translated by using the Edit Sequence tool (Lasergene Version 5.1, July 1999). The translated sequences were then aligned using the online multiple sequence alignment tool, Clustal W ( http://www.ebi.ac.uk/clustalw ). GenBank accession numbers The DNA sequences of the partial spa gene from the representative isolates have been deposited in the DDBJ/EMBL/GenBank database under accession numbers EF562604-EF562613 for isolates spa 1- spa 10, respectively. Results Phage typing Of the 125 MRSA isolates, 65 (52%) were typed into four different phage types by the international set of phages. Thirty-four (27.2%) were typed by phages 52/54/85/81, 15 (12%) by phage 81, 10 (8%) by phage 54 and six (4.8%) by phages 54/75. We were unable to categorize the remaining 60 (48%) isolates by any phage of the international set of basic phages. Detection of mec A gene by PCR The DNA fragment of 310 bp was amplified from all the strains that were detected positive for resistance to methicillin by the agar screening method. PCR-RFLP pattern of spa gene All the 125 MRSA isolates were PCR positive for spa gene. The sizes of the PCR products ranged from 1150 bp to1420bp [Figure - 1]. These PCR products, after digestion with BSP 143 II restriction enzyme, showed five distinct spa banding patterns [Figure - 2]. The types of PCR-RFLP patterns of MRSA isolates are shown in [Table - 1]. Among the 125 isolates, pattern I was most common (34.4%) followed by pattern II (24.8%), pattern V (17.6%), pattern IV (13.6%) and pattern III (9.6%), respectively. The smallest fragment of 243 bp remained same in all patterns while the middle-sized fragment was either 264 bp (in patterns II, III and IV) or 438 bp (in patterns I and V). The high molecular weight fragment was of different sizes in all the patterns, i.e., 715 bp for pattern I, 667 bp for pattern II, 643 bp for pattern III, 811 bp for pattern IV and 739 bp for pattern V. Sequence analysis of spa gene The 1150-1420 bp PCR-amplified fragments of the spa gene from all five PCR-RFLP patterns from 2-3 isolates were sequenced. All the sequences showed two restriction sites for BSP 143 II enzyme. Whereas the first restriction site was fixed at 243 bp in all the sequences, the position of the second restriction site varied. It was either at the 507 bp (patterns II, III and IV) or at the 681 bp (patterns I and V) position. All the five patterns of the spa gene had a polymorphic X region that contained a variable number of 24 bp tandem repeats. The organizations of these repeats in the polymorphic X region of the spa gene of each isolate and their corresponding amino acid alignments are shown in [Figure - 3]. The individual spa repeat with nucleotide and its deduced amino acid sequences are shown in [Figure - 4]. These repeats have variable sequences and are assigned specific letter codes according to the literature. [16] Each type of PCR-RFLP pattern ( spa type) thus had a mixture of different combinations of tandem repeats [Figure - 5]. Isolates with pattern III were found to contain a minimum number (six) of repeats whereas isolates with pattern IV contained the maximum number (13) of repeats. Patterns II, I and V were found to contain 7, 9 and 10 repeats, respectively [Figure - 5]. Upstream to the 24 tandem repeats, the sequences also had a tandem repeat of 174 bp with variable copy numbers: 3.5 in patterns II and IV, 3.9 in pattern III and 4.5 in patterns I and V. Apart from the non-synonymous mutations, some synonymous mutations were also observed in the 24 bp repeats resulting in nucleotide substitution with no amino acid change [Figure - 3]. Discussion Strain typing is an integral part of epidemiological surveillance and infection control in hospitals. For over 20 years, the National Staphylococcal Phage Typing Center in India has been using bacteriophage typing as its S. aureus strain typing method. However, recently, the value of phage typing was impaired by the poor typeability of MRSA strains and thus necessitated the implementation of more efficient and advanced methods for strain typing. [7] We explored molecular methods for typing MRSA isolates. Among various methods, we selected spa genotyping as it is simple, rapid and practical to monitor variations in MRSA populations from divergent regions. [16] Variations in the spa gene sequences have been the basis for PCR typing of S. aureus . It has shown a good correlation between the variable number of repeats, epidemicity and phage type. [16],[17],[18] Furthermore, we also investigated the possibility of the usefulness of the PCR-RFLP patterns of the spa gene in differentiating the MRSA strains, which are otherwise non-typeable by the phages. We were able to type all the 125 MRSA isolates into five different groups by PCR-RFLP [Figure - 2]. This is contrary to the routine phage-typing method where only 52% ( n = 125) of the isolates were typeable while the rest of them remained non typeable. Among these 60 (48%) non phage typeable isolates, four different PCR-RFLP patterns were observed, i.e., pattern I (11 isolates), pattern II (10 isolates), pattern IV (17 isolates) and pattern V (22 isolates), with no isolates falling in pattern III. Further, RFLP patterns IV and V were seen only among the non phage typeable isolates while pattern III was seen only in the phage typeable strains [Table - 1]. In a study by Schmitz et al., [17] 183 MRSA isolates were categorized by a protein A gene-based PCR method, which were otherwise showing identical PFGE patterns. We have also observed heterogeneity within various phage-typed strains. As 34 isolates belong to phage type 52/54/85/81 and 15 isolates belong to phage type 81, each group could be further categorized into three different RFLP patterns, i.e. types I, II and III. Similarly, 10 isolates belonging to phage type 54 were further subgrouped into two patterns of types I and II. The least number of six isolates belonging to phage type 54/75 had only one type (type II) of RFLP pattern [Table - 1]. Sequencing of the PCR products for spa gene fragments from various isolates representing different categories also confirmed the RFLP pattern. The nucleotide sequence of isolates with different PCR-RFLP patterns showed a variable number of 24 bp tandem repeats (6-13 in number). RFLP pattern III with a minimum number of repeats, i.e. six repeats, was seen only in phage typeable strains typed either with phage 81 only or in combination of phage 81 with other phages and showing an extended spectrum of phage type. RFLP pattern II with seven number of tandem repeats was seen in isolates of phage type 54 and phage type 54/75 only. RFLP patterns IV and V with 13 and 10 number of repeats respectively were seen only in non phage typeable strains.This indicates that there is a correlation between phage type and number of tandem repeats. Other workers have also reported that the discrimination of MRSA isolates is possible by determining the repeat sequence numbers within the X region of the spa gene. [10],[18] Van Belkam et al. [10] reported that S. aureus strains of the same lysogroup have different number of repeats in the spa region. These 24 bp repeats have been assigned letter codes according to the literature. [17] Organization of these repeats in the polymorphic X region of the spa gene of each isolate is represented as a spa type repeat code. These repeat codes were used for bacterial typing by Shopsin et al. [16] and Larry Koreen et al. [19] We have observed three new spa repeat patterns (patterns I, III and IV) among our isolates that have not been reported so far from anywhere else [Figure - 5]. spa -II and spa -III have a similar repeat pattern except that spa -III has the last Q1 repeat missing. One partial repeat of 15 nucleotides (AAAGAAGACGGC/TAAC) was observed at the 3′ of all spa type repeat codes in the X region. From the repeat profile, we categorized 16 repeats at the nucleotide level and only six at the amino acid level [Figure - 3]. This difference is due to the emergence of synonymous mutations to conserve the amino acid sequence under different levels of evolutionary pressure. Some non-synonymous mutations were also observed that lead to the change in the amino acid at the fourth, seventh and eighth positions in eight amino acid tandem repeats. These repeats had a G substitution at the seventh and a K at the eighth position and pattern I had a unique substitution of Q at the fourth amino acid in some of the repeats [Figure - 3]. These mutations may have evolved under a high level of evolutionary pressure. [16] The length polymorphism in the gene was not only due to the 24 bp tandem repeats but also involved the 174bp tandem repeats because one complete 174bp repeat was missing in patterns II, III and IV. [20] The copy number of this 174 bp repeat varied among different patterns. These repeats are located near the 5′ terminal of the spa gene, which is upstream to the 24 bp tandem repeats. The encoded peptide region of the protein containing the 174bp repeats interacts with the host IgG. [20] Therefore, this variation is important from the point of view of the host-parasite interaction as it can allow the bacteria to evade the host immune system. On BLAST search ( http://www.ncbi.nlm.nih.gov/BLAST/ ), sequences from patterns II, III and IV showed maximum similarity to S. aureus V8 strain from Welcome Laboratory while the rest of the patterns I and V showed maximum similarity to S. aureus subspecies aureus COL. The study demonstrated that the PCR-RFLP pattern analysis of the spa gene could further discriminate the MRSA strains that were otherwise indistinguishable by an international set of basic phages. We were able to differentiate the non phage typeable (48%) MRSA isolates by PCR-RFLP into four different groups. We also report here three new spa repeat patterns among MRSA strains. DNA sequence analysis of the amplified spa gene fragment of the representative RFLP patterns indicates towards a correlation between the number of tandem repeats and the phage type; however, there is need for further validation on a large number of strains. References
Copyright 2009 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb09037f2.jpg] [mb09037t1.jpg] [mb09037f5.jpg] [mb09037f3.jpg] [mb09037f4.jpg] [mb09037f1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}