
|
Indian Journal of Medical Microbiology
Medknow Publications on behalf of Indian Association of Medical Microbiology
ISSN: 0255-0857 EISSN: 1998-3646
Vol. 27, Num. 3, 2009, pp. 191-201
|
Indian Journal of Medical Microbiology, Vol. 27, No. 3, July-September, 2009, pp. 191-201
Review Article
Role of Borna Disease Virus in neuropsychiatric illnesses : Are we inching closer ?
Thakur R, Sarma S, Sharma B
Department of Microbiology, IHBAS, Dilshad Garden, Delhi-110 095
Correspondence Address:Department of Microbiology, IHBAS, Dilshad Garden, Delhi-110 095
drrajeevthakur@rediffmail.com
Date of Submission: 03-Mar-2008
Date of Acceptance: 19-Apr-2008
Code Number: mb09061
PMID: 19584498
DOI: 10.4103/0255-0857.53200
Abstract
The biological cause of psychiatric illnesses continues to be under intense scrutiny. Among the various neurotropic viruses, Borna disease virus (BDV) is another virus that preferentially targets the neurons of the limbic system and has been shown to be associated with behavioural abnormalities. Presence of various BDV markers, including viral RNA, in patients with affective and mood disorders have triggered ongoing debate worldwide regarding its aetiopathogenic relationship. This article analyses its current state of knowledge and recent advances in diagnosis in order to prove or refute the association of BDV in causation of human neuropsychiatric disorders. This emerging viral causative association of behavioural disorders, which seems to be inching closer, has implication not only for a paradigm shift in the treatment and management of neuropsychiatric illnesses but also has an important impact on the public health systems.
Keywords: Borna disease virus, emerging infection, neuropsychiatric illnesses
The aetiology of psychosis is considered to be multifactorial. While genetic and behavioural components of psychosis have been the focus of intense study, infection as an aetiological factor has received little attention. Persistent viral infection in the central nervous system (CNS) can induce progressive neurological damage with diverse pathological manifestations. However, a direct linkage between viral infection and neurobehavioral disturbances associated with human psychiatric disorders has not been identified. Psychopharmacological studies have attributed these disorders to be associated with neurotransmitter imbalances. Although various viral pathogens like Cytomegalovirus, Herpes simplex virus and Influenza virus have been shown to be associated with psychiatric disorders, [1],[2],[3] their conclusive role as causative agents of such disorders have never been established. Of late, the Borna disease virus (BDV) is one more agent added to the list and is under close scrutiny as the causative agent of neuropsychiatric disorders.
BDV is the causative agent of Borna disease (BD), a fatal non-purulent meningoencephalitis that was originally described in Germany and Switzerland. It is named after the town Borna in Sexonomy/Germany, where an epidemic attack of BD occurred in cavalry horses 100 years ago. [4] Knowledge about disease in animals has initiated research on its association with human psychiatric disorders. Gaps in understanding BDV pathogenesis in humans have triggered controversies regarding its aetiopathogenic relationship. Despite encouraging results of earlier studies, the issues of clinical significance and the establishment of human infection are still being debated.
Epidemiology Not much is known regarding the epidemiology of BDV. Originally, it was supposed to be a natural pathogen of horses and sheep in South Eastern Germany, but now it is known to infect several species ranging from birds to primates across the world. [5] Wild birds may act as subclinically infected carriers of BDV, which can explain its seasonal occurrence (spring and early summers) and higher prevalence in some years compared with others. However, their exact role in the epidemiology of BDV remains mysterious. [6] BD is supposed to be a zoonotic disease. Its incubation period varies from a few weeks to many months. The virus is shed with secretions from the respiratory tract, conjunctiva and faeces of infected animals [7] and is supposed to spread intranasally. [8] BDV has a particular predilection for the neurons of the limbic system, basal ganglia and the brain stem and diffuses throughout the CNS. It replicates locally in the neurons and migrates intra-axonally and spreads by direct cell to cell contact via neuroreceptors in the olfactory epithelium to the adjacent brain tissue. [9]
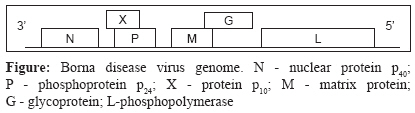
Genomic Organization BDV is an enveloped, non-cytolytic, non-segmented negative strand RNA virus of size 8.5-10.5kb, belonging to family Bornaviradae of the order Monnegaviradae. The other viruses in the Monnegaviradae group are Filoviradae, Paramyxoviradae and Rhabdoviradae. [5] BDV is a spherical enveloped virus with a diameter of 90-130nm. [10] The virus particle contains an internal electron-dense core (50-60nm) and a limiting outer member envelope covered with spikes (7nm long). The virions are assembled at the cell surface from where only a small number of virions are released by budding. [11] It replicates and transcribes its genome in the nucleus and uses cellular RNA splicing machinery to regulate viral gene expression. [12] The BDV genome [Figure - 1] comprises of six open reading frames (ORFs): ORF I codes for the nucleoprotein p 40 (NP), ORF II for phosphoprotein p 24 (P), ORF III for matrix protein p 16 (M), ORF IV for glycoprotein (G) and ORF V for p 180/190 , the phosphopolymerase (L). [13] A further ORF overlaps the P gene and encodes the protein p 10 (X). [14] The phosphoprotein P of BDV is an essential cofactor of the viral RNA-dependent RNA polymerase and its cofactor activity is negatively regulated by phosphorylation unlike other viral P proteins. [15] The matrix protein M of BDV is an integral component of the viral ribonucleoprotein (RNP) complex without interfering with the viral polymerase activity. This is a unique feature among negative-strand RNA viruses and therefore BDV-M is regarded as a pre-requisite for the establishment of BDV persistence. [16] The X is a non-structural protein and can interact with viral polymerase cofactor P thus regulating polymerase activity. [17] The lack of X in virus particles may facilitate polymerase activity early in infection, but the presence of X in persistently infected cells may result in partial inhibition of the polymerase thus contributing to viral persistence. [18]
BDV Life Cycle BDV entry into the cell occurs by receptor-mediated endocytosis. [19] BDV attaches to the host cells via the surface glycoprotein (G) as antibodies to G have been demonstrated to have a neutralizing activity. [20] The N-terminal part of the BDV glycoprotein is sufficient for receptor recognition and virus entry. [21] The N-terminal of gp43 is involved in the pH-dependent fusion after internalization of BDV by receptor-mediated endocytosis. [19],[21] This fusion event releases the BDV RNP, which is then transported to the cell nucleus where BDV transcription and replication occur. The RNA-dependent RNA polymerase (L) of BDV is postulated to be the catalytic enzyme of replication and transcription. BDV polypeptides, with the exception of the N-terminal part of the glycoprotein, are translated from polycistronic mRNAs, a unique situation among mononegaviruses. However, the mechanism involved in the control and trafficking of viral RNP across the nuclear export remains largely unknown. The assembly and release of BDV are poorly understood. Thin sections of BDV-infected cells show the presence of intracytoplasmic virus-like particles with morphologic characteristics similar to those described for partially purified cell-free BDV infectious particles. These particles show no association with the cisternae of the endoplasmic reticulum, Golgi complex or other intracytoplasmic membrane. The assembly process and site of virus maturation have not been identified. Budding of BDV particles is only observed at the cell surface of BDV-infected Madin Darby Canine Kidney cells after treatment with n-butyrate. [22]
Pathogenesis
The CNS is unique in its response to viral infection. There is lack of specialized lymphatic drainage, which potentially limits and delays viral antigen recognition. Persistence of BDV in the CNS may be supported by the blood brain barrier, paucity of immune elements and non-cytolytic strategy for viral replication. [23] BDV is neurotropic and infects mainly neurons in its natural host. It has a high affinity for the limbic system, which is involved in the regulation of behaviour, memory and emotion. BD is a progressive immune-mediated disease. The virus itself is not involved in the destruction of the brain tissue but damage may occur indirectly by triggering of the cell-mediated immune response. [24] Both CD 4 and CD 8 T cells are present in the CNS and their elimination or functional blocking prevents both BDV infection-induced neurological symptoms and histological changes in the brain. [25] In mice, BDV-specific CD 8 T cells have been demonstrated in the brain but not in the peripheral lymphoid organs. [26] CD 8+ T cells exert effector cell functions and their activity results in the destruction of virus-infected cells. Thus, the cytotoxic T-cell response determines whether an ensuing immunopathological disease is controlled or a persistent infection is established. Polymorphonuclear leucocytes can harbour viruses although it has rarely been shown that viruses infect these cells directly. On the other hand, it has been demonstrated that granulocytes take up virions and are capable of phagocytosing infected cells or cell debris.
Predominant lesions found in the CNS consist of perivascular infiltrates of lymphocytes, plasma cells and other mononuclear cells. In the nuclei of the infected neurons, aggregates of virus-specific material form the Joest Degan inclusion bodies , characteristic of BDV infection. [27] The virus propagates in the form of RNP as virus particles are not detected in the astrocytes, oligodendrocytes, ependymal cells and Schwann cells in the peripheral nervous system. [28] Neutralizing antibodies appear late during infection and have been implicated in restricting viral replication in the nervous system. [29] Increased levels of interleukin (IL)-6, tumour necrosis factor-α, IL-α and inducible nitric oxide synthase mRNA in BDV-infected rat brain correlates with the degree of inflammation and the severity of neurological signs. [30]
The onset of clinical signs coincides with the appearance of an inflammatory reaction in the brain that reaches its maximum severity in 30-40 days after infection. [28] This extensive inflammatory reaction leads to neuronal destruction that in some cases may progress to hydrocephalus. Pharmacological studies indicate that behavioural disturbances associated with BD might be due to BDV-induced alteration of the dopamine system. [31] Decreased numbers of D2 and D4 receptor binding sites are found in the nucleus. Besides dopamine, alteration involvement of other neurotransmitter systems have been supported by changes in the mRNA levels of cholecystokinin, glutamic acid decarboxylase and somatostatin. [32] These pharmacological abnormalities are likely consequences of an inflammatory response. However, cortical cholinergic denervation was observed in BDV-infected rats in the absence of any significant cell destruction. [33]
Diagnosis of BDV Infection
As on date, there is no conclusive evidence that BDV is the causative factor of human diseases. For almost a decade antibodies to BDV components were the only markers suggesting infection. These antibodies were found not only in patients with neuropsychiatric diseases but also in immunocompromised as well as healthy individuals. Thus, the need to look for other markers of the infection became very much pertinent.
For diagnosis of BDV infection in human specimens and understanding its aetiological role, detection of proteins that are expressed during infection has become a critical area of investigation. BDV antigen and nucleic acid has been detected in peripheral blood mononuclear cells (PBMCs) from psychiatric patients by novel techniques. PBMC from severely depressed patients subsequently served as the source of the first BDV isolate from humans by long-term cocultivation procedures. [34] A putative human isolate of BDV was also obtained from granulocytes [35] and the first human brain isolate from a schizophrenic patient was reported in 2000. [36] Various modalities have been used for the diagnosis of BDV infection. These include detection of virus-specific antibodies in blood, isolation of virus-specific RNA from PBMCs and brain, detection of circulating plasma antigens and circulating immune complexes (CICs) and T cell proliferation assay.
Detection of virus-specific antibodies in blood
BDV infection in humans was first detected by serological methods, mainly immunofluorescence assays (IFA). IFA with fixed antigen accumulating in the nucleus of infected cells became an indispensable tool for monitoring and titrating antibodies in the mid 1980s. [37] Complement fixation and Ouchterlony tests, which measured either antibodies or precipitated antigen, were additionally helpful, but the sensitivity of these tools always remained questionable. [38]
At the present moment, immunofluorescent (IF)-based assays are commonly used by most research laboratories to detect antibodies against BDV infection. All IF-based assays utilize recombinant products (N, P or M) to measure antibodies. [39],[40] Modification of IFA with monoclonal anti-N antibodies in a double staining protocol was used to improve specificity [37] and focus immunoassays and cell enzyme linked immunosorbent assay (ELISA) were developed to enable faster monitoring of antibodies against BDV. [41] However, all these assays were time consuming and their endpoint titration was subjective. Electrochemiluminescence immunoassays (ECLIA) enabled the amount of antibodies to be determined more precisely but they did not provide any new insights into human BDV infection. [42],[43]
Based on a study by Allmang et al. using modified IFA involving the use of 3M urea, it was reported that human anti-BDV antibodies were of low avidity compared with animal antibodies thus raising doubts about the specificity of human antibodies to BDV. [44] A later avidity experiment with 6M urea, tested by ELISA with native antigen in a double sandwich format, revealed low- and high-avidity antibodies in both species, consistent with findings in other persistent virus infections. [45]
The drawback of all of these recombinant ELISAs was that they had a low sensitivity for BDV antibodies, which might explain the heterogeneity of data from different studies. This low sensitivity can be explained by the fact that native conformational epitopes are more efficient targets to detect anti-BDV antibodies than recombinant proteins used in these assays.
ELISA developed by Bode et al. is based on a double sandwich format using antibody-stabilized monoclonal antibodies that recognize conformational epitopes of BDV N (p40) and P (p24) proteins and these epitopes recognize immunoglobulin (Ig) G in the test serum. This assay thus represents the method of choice for detecting BDV-specific antibodies as it is simple, quantitative, highly sensitive and specific and thus may be labelled as a gold standard test. [46],[47]
Western blots (WBs) serve as confirmatory tests in the diagnosis of many viral infections and have been useful in identifying antibodies against the N and P proteins in serum and cerebro spinal fluid (CSF), but they have not come to the forefront of human BDV diagnosis as they have been shown to be less sensitive than the newer generation ELISAs. [34],[46]
Several studies based on antibody detection have demonstrated a relation between BDV and neuropsychiatric manifestations [Table - 1]. Results of antibody studies by IFA vary between 0 and 16% in psychiatric patients, 0 and 51% in healthy controls and 0% in blood donors, whereas by ELISA they lie between 0 and 24%, 0 and 1% and 0%, respectively. WB results vary between 6.5 and 45% in psychiatric patients, 0 and 1% in healthy controls and 4.7 and 9% in blood donors.
The data using these serological tests employing recombinant antigens do not suggest an association of BDV or a related agent with specific psychiatric diseases in human beings but a newer format of ELISA developed by Bode et al. using native antigens have shown a much higher level of antibodies in psychiatric patients (71.4%) than in healthy subjects (20.3%). [46] It can thus be used as an indicator of infection. However, results have been shown to vary within different geographical regions due to a difference in the methods used for the detection of the antibodies.
Isolation of virus and virus-specific RNA from PBMCs and brain
- Demonstration of BDV RNA in PBMCs and granulocytes.
1.1 BDV RNA in PBMCs of psychiatric patients was first demonstrated by Bode et al . in the mid 1990s by nested reverse transcription polymerase chain reaction (RT-PCR). [34],[48] Thereafter, many workers isolated the BDV-specific RNA from PBMCs of healthy and diseased humans by using RT-PCR in lysates of PBMC isolates from blood after Ficoll gradient separation. Despite the fact that granulocytes represent potential contaminants of PBMC, [35] the reported isolation of Bona virus from granulocytes focused the search for BDV-specific RNA on this fraction of white blood cells. [35],[40] The above results could be explained by the ability of granulocytes to incorporate antigen-antibody complexes and possibly RNPs floating in the blood.
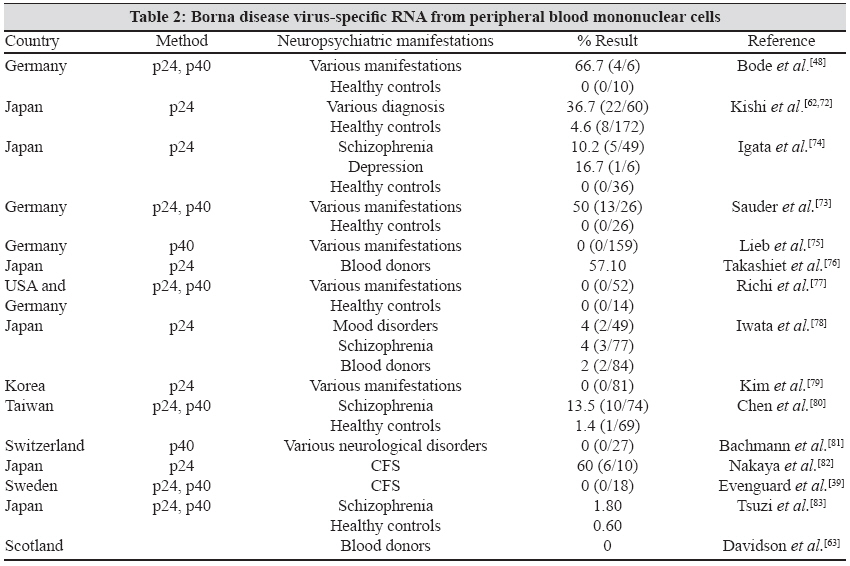
1.2. Various studies were conducted all over the world to detect a relation between BDV-specific RNA in PBMC and its relation to neuropsychiatric manifestation [Table - 2].
1.3. The prevalence of BDV-derived nucleic acid (BDV RNA) has been shown to vary between 0 and 60% in neuropsychiatric diseases, 0 and 57% in the healthy population and 0 and 57.1% in the average blood donors. Most researches have reported a higher prevalence of RNA-positive PBMCs in psychiatric patients than in healthy people, but the positive rates differed among laboratories. Despite some reports on the absence of BDV RNA in psychiatric patients, [41],[47] the occurrence of BDV RNA in human samples is generally accepted and is supported by the finding of BDV RNA in brain samples taken at autopsy. - Isolation of virus and virus-specific RNA from the brain.
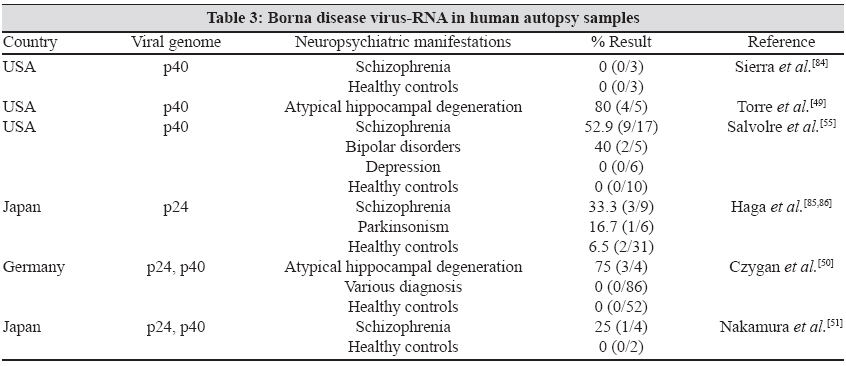
2.1. BDV-specific RNA has been isolated from brain samples at autopsy from patients with various mental disorders. BDV-derived nucleic acid in brain samples were detected by means of nested RT-PCR, in situ hybridization and immunohistochemistry. [49] In these studies, the p24, p40 and first intergenic region were almost identical to the corresponding regions of the animal strain of He/80. [50] All research groups are aware of the possibility that the finding of BDV RNA might indicate contamination of human and animal samples, but this potential has been minimized by using improved safety conditions. Overall, arguments questioning human BDV infection based on contamination of human and animal samples have gradually faded. [40]
2.2. In a study conducted in the USA, the genomic sequence of BDV-RNA isolated from the brain of three patients was identical to a virus that originated in a German horse. Thus, it seems that BDV variants are wide spread and are capable of infecting both horses and humans. It also explains the extraordinary stable nature of the BDV genome in different geographical regions. [50] In a study, viral RNA was detected by RT-PCR and in situ hybridization in the neurons of three of 12 brain regions. There was associated lymphocytic infiltration, which was not detected in the BDV-negative brains. [4] BDV was also recovered after intracerebral inoculation of brain extract into new born gerbil. Nucleotide sequences of this viral isolate differ only by 2% from the standard laboratory strain. [51] Various studies were conducted all over the world to isolate BDV-RNA from the brain [Table - 3]. According to these studies, the rate of isolation of BDV-RNA from the human brain varied between 0 and 80% in patients with various neuropsychiatric manifestations and 0 and 65% in the healthy control group. The results of isolation depended on the sensitivity of the assay as well as on the post-mortem interval. A short post-mortem interval had a higher chance of isolation.
Detection of circulating plasma antigens and CICs
The assay to capture plasma antigen represents a breakthrough in the diagnosis of BDV infection due to the fact that although there may be a low viral load during initial infection, free antigens start appearing in the body fluids quite early. Large amounts of plasma antigens were detected in a cohort of severely depressed patients by ELISA and the severity of symptoms appeared to correlate with the concentration and duration of antigenaemia. [46] In contrast, the presence of antigen in plasma occurs rarely and at low levels in healthy individuals. Thus, plasma antigenaemia indicates an acute, transient and productive phase of infection. During a productive phase, the appearances of plasma antigen parallel the disease process until antibodies bind the antigens to from CICs. These complexes are measurable for weeks or months in patients followed longitudinally. Changes in the relative amounts of CIC and plasma antibodies over time can be used as a prognostic marker. CICs represent the prevailing infection marker and are found in more than 90% of the patients with major depressive disorders or bipolar disorders during acute depression. [45] Screening blood samples solely for CICs may be sufficient to confirm BDV infection. [47] Moreover, the frequency and relative stability of BDV CICs make them currently the best available screening marker of BDV infection.
The BDV specificity of CICs was demonstrated in WBs after precipitation with polyethylene glycol or isolation on protein G-Sepharose columns. The immune complexes consist of N and P monomers or dimers and most probably N-P heteromers bound to IgG antibodies and are very stable, like the antigens. Molecular studies on antigen-antigen interactions and their biochemical characterization are ongoing.
T cell proliferation assay
The T cell proliferative response has also been used as an indicator of BDV infection. Positive proliferative responses to both BDV p40 and p24 proteins were detected in 9% of the patients with mood disorders, 4% of schizophrenic patients and 2% of blood donors. [42]
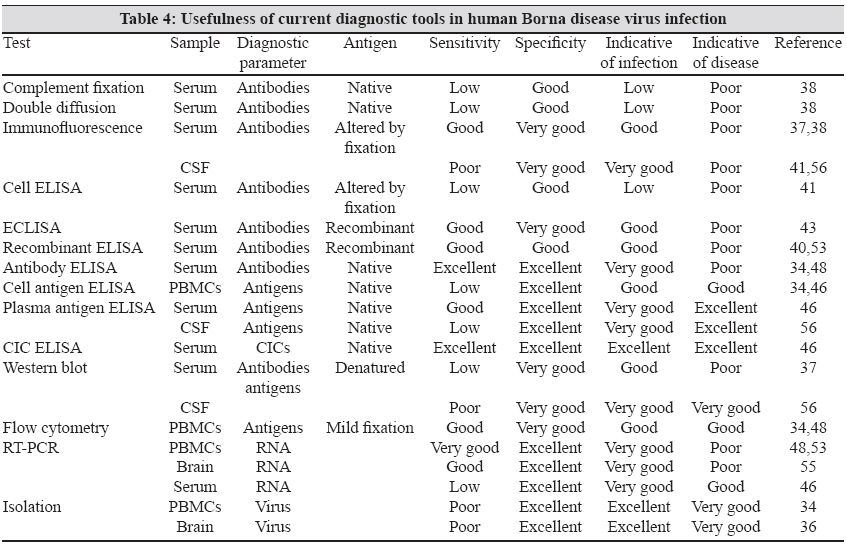
Many recent studies on BDV infection have reported the use of triple ELISA, [45] which measures CIC, plasma antigen and antibodies in the same serum sample as combined measurement of antibodies and CIC in serum enhances the sensitivity of the detection system. The usefulness of various diagnostic tools in human BDV infection has been summarized in [Table - 4]. Among the various tests, CIC ELISA and virus isolation from PBMCs/brain constitute the best markers, indicative of infection. The best indicators of viral disease are antigen ELISA and CIC ELISA, both of which also correlate well with severity of viral disease.
Role of BDV in Health and Disease
As defined in the "Diagnostic and Statistical Manual of Mental Disorders of the American Psychiatric Association", 4 th Ed. (DSM IV), half of the mood disorders are thought to be of an "endogenous" origin with unknown causes, probably driven by an imbalance in the neurotransmitter network. Serious conditions like bipolar disorders and major depressive disorders are present in 1-5% of the population throughout life following a relapsing episodic course. Thus, some one hundred million people worldwide suffer chronically from alternating between mania and depression or from recurrent depressive episodes. [52]
BDV infections have been linked with various psychiatric disorders like major affective disorders, schizophrenia, obsessive compulsive disorder (OCD) and chronic fatigue syndrome by various groups of workers. [37],[40],[47] In affective disorders, depending on the time point and the diagnostic parameters used, the point prevalence of BDV has been found to vary from 2 to 100%. This wide range of positivity was due to the fact that during the well state the positivity was found to be low whereas during episodes of acute depression the positivity in the same patients was found to be increased. Therefore, most of the subsequent clinical studies were designed as longitudinal investigations. [45]
Among patients in the acute stage, 40-50% had BDV antigen-positive PBMCs in the group of severe cases [53],[54],[55] and the probability of virus isolation increased with an increase in the severity of diseases. [34] In a study performed by Bode et al. in 2001, most of the acutely depressed major depressive disorder and bipolar disorder patients were CIC positive (>90%), and the severity of symptoms correlated with high plasma levels of CIC and antigen. Among patients of all clinical categories of depression in the study, about 50% were CIC positive. [46]
In a study performed by Bode et al. in 2003, 40 OCD patients were followed longitudinally and a considerable proposition was found to express BDV infection markers (CICs, plasma antigens or antibodies) in their blood. The duration and levels of antigenaemia and CICs paralleled severe long lasting courses of this disorder, indicating a chronically productive infection state. However, in classical major depressive disorder and bipolar disorder patients, the periodicity of antigenaemia was predominant, with a peak during clinical episodes. [45]
In chronic fatigue syndrome, BDV prevalence based on detection of RNA ranges from 0% in Swedish patients [46] to> 80% in Japanese family clusters. [53] BDV RNA was also positive in a patient analysed by an Austrian group. [54] BDV was isolated from an American chronic fatigue syndrome patient [47] and activated infection (CICs and plasma antigen) was detected in one-thirds of the patients suffering from chronic fatigue syndrome, indicating that the virus persists and long-term antiviral treatment may be beneficial to ameliorate symptoms. [45]
The presence and expression of BDV in human brain have been a matter of debate. Also, the presence of BDV RNA in the brain does not necessarily reflect an active state of viral replication. In samples obtained at autopsy from American and European brain banks, PCR amplifiable BDV-specific RNA (found in 53 and 40% of the samples, respectively) was derived only from patients with schizophrenia and bipolar disorder and not from cases of neurological disorders. [55]
BDV was isolated from a Japanese schizophrenic patient in 2000 [41] and the only report indicating BDV activity in the human brain correlating with disease was a report by Deuschle et al. in 1998 in which BDV antigens were detected in CSF samples from patients with major depressive disorders but not from other psychiatric disorders. [36],[56]
BDV infections can remain clinically silent in healthy humans. Various studies have demonstrated the presence of BDV infection markers in both healthy controls and blood donors. Several research groups have reported a seroprevalence of 2-3% in healthy controls. [37] A carrier state in up to 5% of the population was reported by detecting BDV RNA in PBMCs. [53]
By IFA, ECLISA and WBs BDV antibodies were found in 0-15%, 0-1% and 0-1% of healthy controls, respectively, [57],[58],[59],[60],[61] whereas by ELISA, 20.3% of the healthy controls were found to be antibody positive using native antigen. [46] CICs were also found to be positive in 26.3% of healthy volunteers. [45] BDV RNA in PBMCs was demonstrated in 0-57% of the healthy population and BDV RNA from human brains was detected in 0-65% of the healthy control group.
In blood donors, CIC was found in 20-30% of the cases and 2-4% were IFA positive. [45] By IFA, ECLISA and WBs, BDV antibodies were found in 0%, 0% and 4.7-9% respectively by other workers. [42],[62] A prevalence of 33% was reported in Australian blood donors [45] whereas a prevalence of 0% was reported in Scotland blood donors by RT-PCR for BDV-RNA in PBMCs. [63]
Human BDV Strains
Only six isolates have been reported to be recovered from humans till date. The sources were PBMCs from two bipolar and one OCD and one chronic fatigue syndrome patient. [34],[64] The fifth isolate (RW98) from the granulocytes of a schizophrenic patient have been withdrawn as a laboratory strain contamination. [65] The sixth isolate was recovered from the brain of a Japanese schizophrenic patient. [36] Human strains were found to have significant differences from the animal strains [34] in their nucleic acid and nucleotide sequences. Furthermore, all human isolates were sensitive to Amantadine treatment in vitro , in contrast to the laboratory strain. [47]
Treatment
There are as yet no specific treatment modalities for patients of BD. However, few agents have been tried for its treatment. One of them is Amantadine. Amantadine is a non-competitive N methyl D aspirate-type glutamate receptor antagonist that also has binding activity to the nicotinic acetylcholine receptor. [66] It appears to act by increasing the synthesis and release of dopamine and by diminishing its neuronal uptake. It is also thought to have some antiviral activity by which it interferes with the uncoating and release of the viral genome into the host cell. Thus, it not only inhibits the replication of wild type BDV in cell in vitro but also prevents replication of naive cells. [47],[67]
Two studies were performed to detect the efficacy of Amantadine in patients with acute major depressive disorder or bipolar disorders: one in Hanover and the other in Berlin. Patients were treated daily with a mean oral dose of 200mg of Amantadine for 12 weeks and the majority showed a significant and rapid clinical response after an average of 3 weeks. The initial laboratory sign of antiviral efficacy in the treated patients was the disappearance of antigen from plasma and PBMCs. Continuous reduction of plasma antigen levels was demonstrated in the 5-year follow-up of a chronic fatigue syndrome patient who partially improved under treatment with Amantadine and was able to carry out daily activities despite residual fatigue. [45]
Conclusion
The role of BDV in neuropsychiatric diseases still remains open to criticism and discussion in light of the current state of knowledge. As studies have revealed a considerable number of silent infections in humans, the issue of BDV infection and BDV disease should be separated. To establish an aetiopathogenic relationship between BDV and human disease remains the major challenge. In asymptomatic patients, BDV infection is characterized by small amounts of CICs with or without free antibodies whereas there is significant expression of plasma antigens and CICs in acute mood disorders. As sources and routes of infection are still unknown, its aetiological role in CNS diseases continues to remain under scrutiny.
Various factors implicating the strength of association of BDV with neuropsychiatric diseases include:
- Unique features of BDV in terms of a broad host range and its ability to selectively interfere with behaviour and social affiliation, which is consistent with both the episodic nature of diseases and sub clinical persistence in human disorders.
- There is a relative tropism for the limbic system and the excitatory field in the hippocampus that makes it a plausible aetiological agent in schizophrenia. [24],[69]
- Experimental infection of animals with BDV may result in symptoms such as aggression, hyperactivity and behavioural disturbances, apathy or motor symptoms that resemble the core features of human psychiatric disorders such as depression and schizophrenia. [70]
3.1. Temporal relationship of infection marker and disease.
3.2. Presence of antigenaemia and CICs in acute depressive episodes.
3.3. Correlation of high levels of CIC with severity of psychiatric symptoms.
3.4 Virus isolation from PBMCs of severely diseased patients with high antigenaemia.
3.5. Multiple analogies of clinical and other parameters between animal models and infected humans.
3.6. Long-term beneficial effect of Amantadine in depressed patients.
Some of the arguments against the clinical significance of BDV in humans include:
- Lack of BDV RNA in PBMCs of some psychiatric patients.
- Incongruence of BDV-specific amplicons and presence of antibodies.
- Questionable activity of human antibodies to BDV in avidity studies.
- Irreproducibility of Amantadine inhibition effect with laboratory strains.
Unique features of BDV along with its incredible pathogenic flexibility and many evidences implicating its increasing strength of association definitely point towards its causative role in human BDV infections. However, the same needs to be proved in natural human infections before claiming behavioural changes due to neurotransmitter imbalances or due to inflammatory reactions in humans. Presently, we can very well speculate on how BDV might alter the neurotransmitter balance in the brains of maniac, depressed or OCD patients. Unravelling the spectrum of BDV pathogenicity requires special efforts and newer diagnostic approaches to further strengthen its pathogenic correlation. Newer and sensitive molecular techniques like real time PCR and sequencing will be very significant tools for epidemiological studies, for tracing the source of infection, analysing the geographic distribution of BDV and elucidating intraspecies and interspecies transmission. As most of the previous studies have applied cross-sectional design, one sample per person, with clinically undefined times of blood testing employing different diagnostic modalities and no periodic monitoring of the course of disease, there is need for multicentric longitudinal studies in different parts of the world for establishing a definitive association of BDV with neuropsychiatric illnesses. At this moment, one thing which is most certain is that the current evidences employing improved diagnostic tests have brought this aetiopathogenic association between BDV and some human mental disorders much closer.[86]
References
| 1. | Albrecht P, Torrey EF, Boone E, Hicks JT, Daniel N. Raised cytomegalovirus antibody level in cerebrospinal fluid of schizophrenia patients. Lancet 1980;2:769-72. Back to cited text no. 1 |
| 2. | Halonen PE, Rimon R, Arhonka K, Jantti V. Antibody levels to herpes simplex type 1, measles and rubella virus in psychiatric patients. Br J Psychiatry 1974;125:461-5. Back to cited text no. 2 |
| 3. | O'Callaghan, Sham P, Takei N, Glover G, Murray RM. Schizophrenia after prenatal exposure to 1957 A2 influenza epidemic. Lancet 1991;337:1248-50. Back to cited text no. 3 |
| 4. | Staeheli P, Lieb K. Bornavirus and psychiatric disorders-fact or fiction? J Med Microbiol 2001;50:579-81. Back to cited text no. 4 |
| 5. | Schneemann A, Schneider PA, Lamb RA, Lipkin WI. The remarkable coding strategy of Borna virus: A new member of nonsegmented negative strand RNA viruses. Virology 1995;210:1-8. Back to cited text no. 5 |
| 6. | Berg M, Johanasson M, Montell H, Berg AL. Wild bird as a natural reservoir of Borna disease virus. Epidemiol Infect 2001;127:173-8. Back to cited text no. 6 |
| 7. | Webster RG. Influenza virus: Transmission between species and relevance to emergence of the next human pandemic. Arch Virol Suppl 1997;13:105-13. Back to cited text no. 7 |
| 8. | Rott R, Herzog S, Bechter K, Frese K. Borna disease:A possible hazard for human? Arch Virol 1991;118:143-9. Back to cited text no. 8 |
| 9. | Morales JA, Herzog S, Kompter C, Frese K, Rott R. Axonal transport of Borna disease virus along olfactory pathways in spontaneously and experimentally infected rats. Med Microiol Immunol 1988;177:51-68. Back to cited text no. 9 |
| 10. | Zimmermann W, Breter H, Rudolph M, Ludwig H. Borna disease virus: Immunoelectron microscopic characterization of cell free virus and further information about the genome. J Virol 1994;68:6755-8. Back to cited text no. 10 |
| 11. | Kohno T, Goto T, Takasaki T, Morita C, Nakaya T, Ikuta K, et al . Fine structure and morphogenesis of Borna disease virus. J Virol 1999;73:760-6. Back to cited text no. 11 |
| 12. | Lieb K, Staeheli P. Borna disease virus- does it infect humans and cause psychiatric disorders? J Clin Virol 2001;21:119-27. Back to cited text no. 12 |
| 13. | de la Torre JC. Molecular biology of Borna disease virus and persistence. Front Biosci 2002;7:d569-79. Back to cited text no. 13 |
| 14. | Wolffe T, Pfleger R, Wehner T, Reinhaerdt J, Richt JA. A short leucin-rich sequence in the Borna disease virus p10 protein mediates association with the viral phospho and nucleoproteins. J General Virol 2000;81:939-47. Back to cited text no. 14 |
| 15. | Schmid S, Mayer D, Schneider U, Schwemmle M. Functional characterization of the major and minor phosphorylation sites of the P protein of Borna Disease Virus. J Virol 2007;81:5497-507. Back to cited text no. 15 |
| 16. | Chase G, Mayer D, Hildebrand A, Frank R, Hayashi Y, Tomonga K, et al . Borna Disease Virus matrix protein is an integral component of the viral ribonucleoprotein complex that does not interfere with polymerase activity. J Virol 2007;81:743-9. Back to cited text no. 16 |
| 17. | Poenisch M, Wille S, Ackermann A, Staeheli P, Schneider U. The X protein of Borna Disease Virus serves essential functions in the viral multiplication cycle. J Virol 2007;81:7297-9. Back to cited text no. 17 |
| 18. | Schwardt M, Mayer D, Frank R, Schneider U, Eickmann M, Planz O, et al . The negative regulator of Borna Disease Virus polymerase is a non-structural protein. J Gen Virol 2005;86:3163-9. Back to cited text no. 18 |
| 19. | Gonzalez-Dunia D, Cubit B, de la Torre JC. Mechanism of Borna disease virus entry into cells. J Virol 1998;72:783-8. Back to cited text no. 19 |
| 20. | Gonzalez-Dunia D, Cubit B, Grasser FA, de la Torre JC. Characterization of Borna disease virus p56 protein, a surface glycoprotein involved in virus entry. J Virol 1997;71:3208-18. Back to cited text no. 20 |
| 21. | Perez M, Watanabe M, Whitt MA, de la Torre JC. N-terminal domain of Borna disease virus G (p56) protein is sufficient for virus receptor recognition and cell entry. J Virol 2001;75: 7078-85. Back to cited text no. 21 |
| 22. | Compans RW, Melsen LR, de la Torre JC. Virus like particles in MDCK cells persistently infected with Borna disease virus. Virus Res 1994;33:261-8. Back to cited text no. 22 |
| 23. | Ludwig H, Bode L, Gosztonyi G. Borna disease: A persistent virus infection of the central nervous system. Prog Med Virol 1988;35:107-51. Back to cited text no. 23 |
| 24. | Shanker V, Kao M, Hamir A, Sheng H, Koprowaski H, Dietzschold B. Kinetics of virus spread and change in levels of several cytokines mRNA in the brain after intranasal infection of rats with Borna disease virus. J Virol 1992;66:992-8. Back to cited text no. 24 |
| 25. | Bilzer T, Stitz L. Immunopathogenesis of virus affecting the central nervous system. Crit Rev Immunol 1996;16:145-222. Back to cited text no. 25 |
| 26. | Duplan V, Suberbielle E, Napper CE, Joly E, Saoudi A, Gonzalez-Dunia D. Tracking antigen-specific CD8 + T cells in the rat using MHC class 1 multimers. J Immunol Methods 2007;320:30-9. Back to cited text no. 26 |
| 27. | Gosztonyi G, Ludwig H. Borna disease-neuropathology and pathogenesis. Curr Top Micrbiol Immunol 1995;190:39-73. Back to cited text no. 27 |
| 28. | de la Torre JC. Bornavirus and the brain. J Infect Dis 2002;186:S241-7. Back to cited text no. 28 |
| 29. | Stitz L, Noske K, Planz O, Furrer E, Lipkin WI, Bilzer T. A functional role for neutralizing antibodies in Borna disease: Influence on virus tropism outside the central nervous system. J Virol 1998;72:8884-92. Back to cited text no. 29 |
| 30. | Stitz L, Dietzschold B, Carbone KM. Immunopathogenesis of Borna disease. Curr Top Micrbiol Immunol 1995;190:75-92. Back to cited text no. 30 |
| 31. | Solbrig MV, Koof GF, Joye JN, Lipkin WI. A neural substrate of hyperactivity in Borna disease change in brain dopamine receptors. Virology 1996;222:332-7. Back to cited text no. 31 |
| 32. | Lipkin WI, Hatalski CG, Briese T. Neurobiology of Borna disease virus. J Neurovirol 1997;3:S17-20. Back to cited text no. 32 |
| 33. | Gies U, Bilzer T, Stitz L, Statiger JF. Disturbances of cortical cholinergic innervation in Borna disease prior to encephalitis. Brain Pathol 1998;8:39-48. Back to cited text no. 33 |
| 34. | Bode L, Durrwald R, Rantam FA, Ferszt R, Ludwig H. First isolates of infectious human Borna disease virus from patients with mood disorders. Mol Psychiatry 1996;1:200-12. Back to cited text no. 34 |
| 35. | Planz O, Rentzsch C, Batra A, Winkler T, Buttner M, Rziha HJ, Stitz L. Pathogenesis of Borna disease virus: granulocyte fractions of psychiatric patients harbour infectious virus in the absence of anti-viral antibodies. J Virol 1999;73:6251-6. Back to cited text no. 35 |
| 36. | Nakamura Y, Takahashi H, Shoya Y, Nakaya T, Watanabe M, Tomonaga K, et al . Isolation of Borna disease virus from human brain tissue. J Virol 2000;74:4601-11. Back to cited text no. 36 |
| 37. | Bode L. Human infections with Borna disease virus and potential pathogenic implications. Curr Top Microbiol Immunol 1995;190:103-30. Back to cited text no. 37 |
| 38. | Ludwig H, Becht H. Borna disease: A summary of our present knowledge. In: Meulen V.ter, Katz M, editors. Slow virus infections of the central nervous system: investigational approaches to etiology and pathogenesis of these diseases. New York NY; Springer; 1977. p. 75-83. Back to cited text no. 38 |
| 39. | Evenguard B, Briese T, Lindh G, Lee S, Lipkin WI. Absence of Borna disease virus infection in Swedish patients with chronic fatigue syndrome. J Neurovirol 1999;5:495-9. Back to cited text no. 39 |
| 40. | Lipin WI, Hornig M, Briese T. Borna disease virus and neuropsychiatric disease: A reappraisal. Trends Microbiol 2001;9:295-8. Back to cited text no. 40 |
| 41. | Ludwig H, Furuya K, Bode L, Klein N, Durrwald R, Lee DS. Biology and neurobiology of Borna disease viruses (BDV), defined by antibodies, neutralizability and their pathogenic potential. Arch Virol Suppl 1993;7:111-3. Back to cited text no. 41 |
| 42. | Fukuda K, Takahashi K, Iwata Y, Mori N, Gonad K, Ogawa T, et al . Immunological and PCR analysis for Borna disease virus in psychiatric patients and blood donors in Japan. J Clin Microbiol 2001;39:419-29. Back to cited text no. 42 |
| 43. | Yamaguchi K, Sawada T, Naraki T, Igata-Yi R, Shiraki H, Horii Y, et al . Detection of Borna disease virus-reactive antibodies from patients with psychiatric disorders and from horses by electrochemiluminescence immunoassay. Clin Diagn Lab Immunol 1999;6:696-700. Back to cited text no. 43 |
| 44. | Allmang U, Hofer M, Herzog S, Bechter K, Staehell P. Low avidity of human serum antibodies to Borna disease virus antigen questions their diagnostic value. Mol Psychiatry 2001;6:329-33. Back to cited text no. 44 |
| 45. | Bode L, Ludwig H. Borna Disease virus infection: A human mental - health risk. Clin Microbiol Rev 2003;16:534-45. Back to cited text no. 45 |
| 46. | Bode L, Reckwald P, Severus WE, Stoyloff R, Ferszt R, Dietrich DE, et al . Borna disease virus-specific circulating immune complexes, antigenemia, and free antibodies - the key marker triplet determining infection and prevailing in severe mood disorders. Mol Psychiatry 2001;6:481-91. Back to cited text no. 46 |
| 47. | Bode L, Ludwig H. Borna disease virus: A threat for human mental health? In: Smith GL, Irving WL, McCauley JW, Rowlands DJ, editors. New challenges to health: the threat of virus infection. Society for General Microbiology, Cambridge University Press: Cambridge, England; 2001. p. 269-310. Back to cited text no. 47 |
| 48. | Bode L, Zimmermann W, Ferszt R, Steinbach F, Ludwig H. Borna disease virus genome transcribed and expressed in psychiatric patients. Nat Med 1995;1:232-6. Back to cited text no. 48 |
| 49. | De la Torre JC, Gonzalez-Dunia D, Cubitt B, Mallory M, Mueller-Lantzsch N, Grasser FA, et al . Detection of Borna disease virus antigen and RNA in human autopsy brain samples from neuropsychiatric patients. Virology 1996;223:272-82. Back to cited text no. 49 |
| 50. | Czygan M, Hallensleben W, Hofer M, Pollak S, Sauder C, Bilzer T, et al . Borna disease virus in human brains with a rare form of hippocampal degeneration but not in patients with common neuropsychiatric disorders. J Infect Dis 1999;180:1695-9. Back to cited text no. 50 |
| 51. | Nakamura K, Takahashi H, Shoya Y, Nakaya T, Watanabe M, Tomonanga K, et al . Isolation of Borna disease virus from human brain tissue. J Virol 2000;74:4601-11. Back to cited text no. 51 |
| 52. | Godwin FK, Jamison KR. Manic-depressive illness. Oxford University Press: New York, N.Y; 1990. Back to cited text no. 52 |
| 53. | Ikuta K, Ibrahim MS, Kobayashi T, Tomonaga K. Borna disease virus and infection in humans. Front Biosci 2002;7:d470-95. Back to cited text no. 53 |
| 54. | Nowotny N, Kolodziejek J. Demonstration of Borna disease virus nucleic acid in a patient with chronic fatigue syndrome. J Infect Dis 2000;181:1860-2. Back to cited text no. 54 |
| 55. | Salvatore M, Morzunov S, Schwemmle M, Lipkin WI. Borna disease study group. Borna virus in brains of North America and European people with schizophrenia and bipolar disorders. Lancet 1997;349:1813-4. Back to cited text no. 55 |
| 56. | Deuschle M, Bode L, Heuser I, Schmider J, Ludwig H. Borna disease virus protein in cerebrospinal fluid of patients with recurrent depression and multiple sclerosis. Lancet 1998;352:1828-9. Back to cited text no. 56 |
| 57. | Zhen FF, Amsterdam JD, Kao M, Shanker V, Koprowaski H, Dietzschold B. Detection of Borna disease virus reactive antibodies from patients with affective disorders by Western immunoblot technique. J Affect Disord 1993;27:61-8. Back to cited text no. 57 |
| 58. | Waltrip RW 2 nd , Buchanan RW, Summerfelt A, Breier A, Carpenter WT Jr, Bryant NL, et al . Borna disease virus and schizophrenia. Psychiatry Res 1995;56:33-44. Back to cited text no. 58 |
| 59. | Horimoto T, Takahashi H, Sakaguchi M, Horikoshi K, Iritani S, Kazamatsuri H, et al . A reverse type sandwich Enzyme -Linked Immunosorbent Assay for detecting antibodies to Borna disease virus. J Clin Microbiol 1997;35:1661-6. Back to cited text no. 59 |
| 60. | Iwahashi K, Watanabe M, Nakamura K, Suwaki H, Nakaya T, Nakamura Y, et al . Positive and negative syndromes and Borna disease virus infection in schizophrenia. Neuropsychobiology 1998;37:59-64. Back to cited text no. 60 |
| 61. | Rybakowski F, Sawada T, Yamaguchi K, Rajewski A, Rybakowski J. Borna disease virus-reactive antibodies in Polish psychiatric patients. Med Sci Monit 2002;8:CR642-6. Back to cited text no. 61 |
| 62. | Kishi M, Nakaya T, Nakamura Y, Kakinuma M, Takahashi TA, Sekiguchi S, et al . Prevalence of Borna disease virus RNA in peripheral blood mononuclear cells from blood donors. Med Microbiol Immunol 1995;184:135-8. Back to cited text no. 62 |
| 63. | Davidson F, Lycett C, Petrik J, Fazakerley JK. Investigation of frequency of active Borna disease virus infection in Scottish blood donors. Vox Sang 2004;86:148-50. Back to cited text no. 63 |
| 64. | de la Torre JC, Bode L, Durrwald R, Cubitt B, Ludwig H. Sequence characterization of human Borna disease virus. Virus Res 1996;44:33-44. Back to cited text no. 64 |
| 65. | Planz O, Rziha HJ, Stitz L. Genetic relationship of Borna disease virus isolates. Virus Genes 2003;26:25-30. Back to cited text no. 65 |
| 66. | Kornhuber J, Weller M, Schoppmeyer K, Riederer P. Amantadine and memantine are NMDA receptor antagonists with neuroprotective properties. J Neural Transm Suppl 1994;43:91-104. Back to cited text no. 66 |
| 67. | Bode L, Dietrich DE, Stoyloff R, Emrich HM, Ludwig H. Amantidine and human Borna disease virus in vitro and In vivo in an infected patient with bipolar depression. Lancet 1997;349:178-9. Back to cited text no. 67 |
| 68. | Amsterdam JD, Winokur A, Dyson W, Herzog S, Gonzalez F, Rott R, et al . Borna disease virus: A possible etiological factor in human affective disorders? Arch Gen Psychiatry 1985;42:1093-6. Back to cited text no. 68 |
| 69. | Gosztonyi G, Ludwig H. Neurotransmitter receptor and viral neurotropism. Neuropsychiatry Clin 1984;3:107-14. Back to cited text no. 69 |
| 70. | Briese T, Hornig M, Lipkin WI. Borna virus immunopathogenesis in rodents: Model for human neurological diseases. J Neurovirol 1999;5:604-12. Back to cited text no. 70 |
| 71. | Rott R, Herzog S, Fleischer B, Winokur A, Amsterdam J, Dyson W, et al . Detection of serum antibodies to Borna disease virus in patients with psychiatric disorders. Science 1985;228:755-6. Back to cited text no. 71 |
| 72. | Kishi M, Nakaya T, Nakamura Y, Zhong Q, Ikeda K, Senzo M, et al . Demonstration of human Borna disease virus RNA in human peripheral blood mononuclear cells. FEBS Lett 1995;364:293-7. Back to cited text no. 72 |
| 73. | Sauder C, Muller A, Cubitt B, Mayer J, Steinmetz J, Trabert JC, et al . Detection of Borna disease virus (BDV) antibodies and BDV RNA in psychiatric patients: Evidence for high sequence conservation of human blood derived BDV RNA. J Virol 1996;70:7713-24. Back to cited text no. 73 |
| 74. | Igata-Yi R, Yamaguchi K, Yoshiki K, Takemoto S, Yamasaki H, Matsuoka M, et al . Borna disease virus and consumption of raw horse meat. Nat Med 1996;2:948-9. Back to cited text no. 74 |
| 75. | Lieb K, Hallensleben W, Czygan M, Stitz L, Staeheli P. No Borna disease virus specific RNA detected in blood from psychiatric patients in different regions of Germany. Lancet 1997;350:1002. Back to cited text no. 75 |
| 76. | Takahashi H, Nakaya T, Nakamura Y, Asahi S, Onishi Y, Ikebuchi K, et al . Higher prevalence of Borna disease virus infection in blood donors living near thoroughbred horse farms. J Med Virol 1997;52:330-5. Back to cited text no. 76 |
| 77. | Richt JA, Alexender RC, Herzog S, Hooper DC, Kean R, Spitsin S, et al . Failure to detect Borna disease virus infection in peripheral leucocytes from humans with psychiatric disorders. J Neurovirol 1997;3:174-8. Back to cited text no. 77 |
| 78. | Iwata Y, Takahashi K, Peng X, Fukuda K, Ohno K, Ogawa T, et al . Detection and sequence analysis of Borna disease virus p24 RNA from peripheral blood mononuclear cells of patients with mood disorders or schizophrenia and of blood donors. J Virol 1998;72:10044-9. Back to cited text no. 78 |
| 79. | Kim YK, Kim SH, Choi SH, Ko YH, Kim L, Lee MS, et al . Failure to demonstrate Borna disease virus genome in peripheral blood mononuclear cells from psychiatric patients in Korea. J Neurovirol 1999;5:196-9. Back to cited text no. 79 |
| 80. | Chen CH, Chiu YL, Shaw CK, Tsai MT, Hwang AL, Hsiao KJ. Detection of Borna disease virus RNA from peripheral blood cells in schizophrenia patients and mental health workers. Mol Psychiatry 1999;4:556-71. Back to cited text no. 80 |
| 81. | Bachmann S, Caplazi P, Ficher M, Ehernsperger F, Cone RW. Lack of association between Borna disease virus infection and neurological disorders among HIV infected individuals. J Neurovirol 1999;5:190-5. Back to cited text no. 81 |
| 82. | Nakaya T, Takahashi H, Nakamur Y, Kuratsune H, Kitani T, Machii T, et al . Borna disease virus infection in two family clusters of patients with chronic fatigue syndrome. Microbiol Immunol 1999;43:679-89. Back to cited text no. 82 |
| 83. | Tsuji K, Toyomasu K, Imamura Y, Maeda H, Toyoda T. No association of Borna disease virus with psychiatric disorders among patients in Northern Kyushu, Japan. J Med Virol 2000;61:336-40. Back to cited text no. 83 |
| 84. | Sierra-Honigmann AM, Carbone KM, Yolken RH. Polymerase chain reaction (PCR) search for viral nucleic acid sequence in schizophrenia. Br J Psychiatry 1995;166:55-60. Back to cited text no. 84 |
| 85. | Haga S, Motoi Y, Ikeda K; The Japan Bornavirus Study Group. Borna disease virus and neuropsychiatric disorders. Lancet 1997;350:592-3. Back to cited text no. 85 |
| 86. | Haga S, Yoshimura M, Motoi Y, Arima K, Aizawa T, Ikuta K, et al . Detection of Borna disease virus in normal human brain tissue. Brain Res 1997;770:307-9. Back to cited text no. 86 |
Copyright 2009 - Indian Journal of Medical Microbiology
The following images related to this document are available:
Photo images
[mb09061t3.jpg]
[mb09061f1.jpg]
[mb09061t2.jpg]
[mb09061t1.jpg]
[mb09061t4.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}