 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
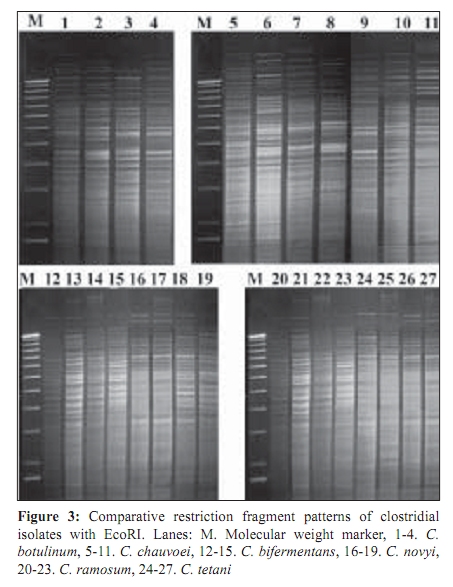
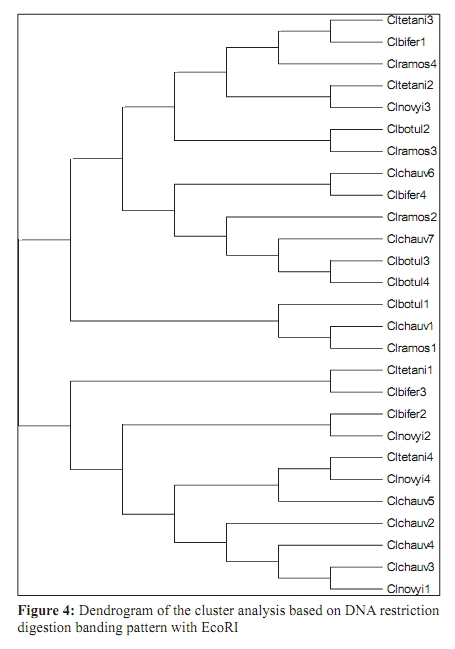
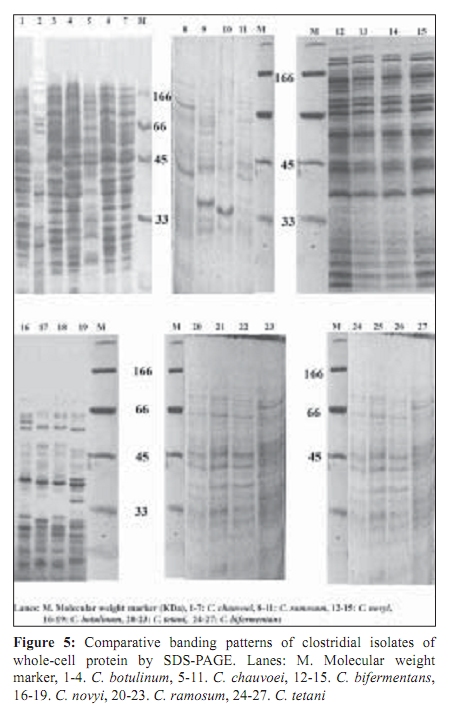
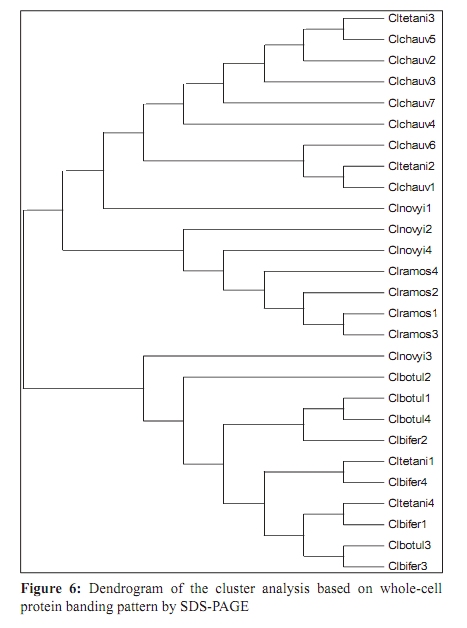
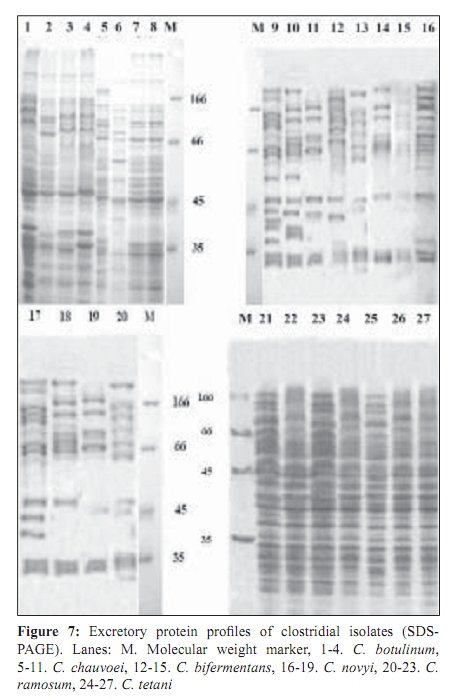
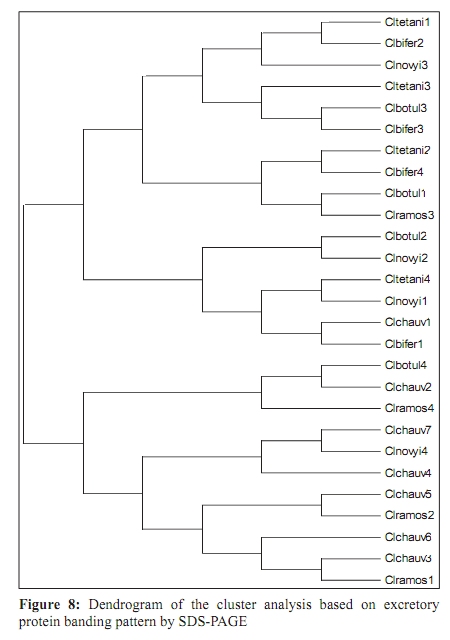
Indian Journal of Medical Microbiology, Vol. 27, No. 4, October-December, 2009, pp. 311-320 Original Article Genetic diversity among toxigenic clostridia isolated from soil, water, meat and associated polluted sites in South India *S Sathish, K Swaminathan Department of Microbial Biotechnology, Bharathiar University, Coimbatore - 641 046, Tamilnadu, India Correspondence Address: *Department of Microbial Biotechnology, Bharathiar University, Coimbatore - 641 046, Tamilnadu, India, sathish3107@gmail.com Date of Submission: 05-Feb-2008 Code Number: mb09090 PMID: 19736399 DOI: 10.4103/0255-0857.55443 Abstract Purpose: To investigate the genetic diversity of toxigenic Clostridium strains isolated from soil, water, meat and its associated polluted sites of Southern India.Materials and Methods: A total of 27 identified isolates of six different toxigenic clostridial species including C. bifermentans , C. botulinum , C. chauvoei , C. ramosum , C. tetani and C. novyi were isolated and characterized by conventional DNA restriction digestion analysis (REA) and by whole-cell and excretory protein patterns on SDS-PAGE. Results: The DNA fragment size ranged from 35-160 kilobases and the protein bands 30-200 KDa, followed by numerical analyses and phylogenetic analyses. Whole-cell protein banding pattern were unique with strains of C. chauvoei , C. novyi and C. ramosum . All the strains were heterogeneous and distinct in restriction digestion pattern and excretory protein patterns. Conclusion: These analyses contribute to the understanding of prevalence of toxigenic clostridial species and phylogeny within the species and assist in development of improved diagnostics and therapeutics for the treatment of clostridial infections. Keywords: Clostridium, phylogenetic analysis, restriction digestion, SDS-PAGE Introduction Clostridia are widely distributed in nature in oxygen-free habitats. Clostridia, in addition to its main soil habitat, were found in dust, sewage, rivers, lakes, sea water, milk, vegetables, fresh meat, fish, insects and the intestinal tract. [1] The genus Clostridium comprises nearly 100 species sub-divided into a majority of non-pathogenic species, 25 to 30 minor pathogens and around 13 classical major pathogens. The notable major pathogens are classified on the basis of lesions and clinical signs into neurotoxic Clostridia ( C. botulinum Groups I, II and III, C. tetani ), histotoxic Clostridia ( C. chauvoei , C. novyi , C. bifermentans , C. septicum C. hemolyticum ) and/or enterotoxic Clostridia ( C. perfringens , C. ramosum , C. difficile , C. septicum , C. sordellii ). Clostridial infections like intra-abdominal infections, pulmonary infections, pelvic infections, brain abscess, skin and soft tissue infections, oral and dental infections, bacteremia and endocarditis are caused by various species such as C. perfringens , C. novyi , C. septicum , C. bifermentans and two species, C. botulinum and C. tetani , that release neurotoxins are responsible for botulism and tetanus, respectively. In addition, recent innovations in food processing and packaging technology have created an increasing interest in the magnitude of botulinal spore contamination of various foods. In the last few years, many researchers have tried to clarify the epidemiology of pathogenic clostridial species and have pointed out the need for a typing scheme. Different methods have been suggested, some of which are already used for other micro-organisms, such as phage typing, serotyping [2] and some of which are based on new techniques, such as autoradiograms of radiolabeled proteins, protein profiles [3] or restriction endonuclease DNA analysis (REA). [4] These methods have proven useful in understanding some aspects of the epidemiology of clostridial species, but no reference scheme has yet been established. The objectives of this study were to study the prevalence of toxigenic clostridia in soil, water, meat and associated polluted sites in Southern India and compare the strains based on DNA restriction pattern and whole cell and excretory protein pattern by SDS-PAGE followed by numerical analyses and phylogenetic analyses. Materials and Methods Sample collection Soil samples (n = 115) were collected from the premises of slaughterhouses, premises of meat-packaging industries, aquatic sediments, river bed, waste dumping areas of meat processing places and commercial meat markets. Soil samples were scooped each at varying depths ranging from 5-20cms. Water samples (n=59) were collected from slaughterhouses, Cochin harbor, lakes, Large-scale meat packaging industries and small water bodies. Water samples from Cochin harbor, (Kerala, India) were collected where large number of dead fishes is thrown back in the water. Water samples from two different slaughterhouses include floor drains, benches where slaughtering of cattle takes place, water discharges, inlet water, storage tanks, immediate carcass wash water, stagnant water from the floor, and sewage. Lakes surrounding the slaughterhouse area were sampled since they are used frequently for disposing meat wastes. Water samples from industries include inlet and immediate wash water, and sewage. Water samples of about five - 30 ml were collected in a sterile collection tubes, sealed, labeled appropriately, brought to laboratory and processed immediately. Meat samples (n is equal to 140) include the carcasses, raw meat and marketed tinned commercial meat. Meat samples of poultry were collected from poultry farm, commercial local butcher's shop as well as from the waste dump. Beef, pork and fish samples were collected separately each from slaughterhouses, local butcher's shop, selling market, meat cleaning and processing area and from waste dump. Meat samples from a meat processing industry include the raw fish, and at several stages of processing and from final processed and packed fishes. Preparation of samples Soil samples (10g) were rinsed with sterile distilled water (100ml) by shaking manually for 3 min. Water samples were serially diluted (1:10) in sterile distilled water. Ten grams of a meat sample was homogenized in a tissue homogenizer with 100 ml PBS for two min. The soil and meat samples were pre-treated for the selection of spore-formers with 70% ethanol (1v/1v) and incubated for 30 minutes or heat-treatment at 80ºC for 10 minutes was used. The pre-treated soil and meat samples were serially diluted (1:10) in phosphate buffered saline. The culture broth and reagents were prepared by pre-reduced anaerobically sterilized technique (PRAS) technique. [5] The pre-treated soil and meat samples and untreated water samples of about 0.5 ml from each dilution were inoculated into 30 ml of pre-reduced thioglycolate broth. The bottles were labeled and incubated at 35ºC for up to 10 days. The culture broth was periodically plated on Clostridial Agar (HiMedia laboratories, Mumbai, India) by spread plate technique and incubated in anaerobic jar with a gas generating kit (HiMedia laboratories, Mumbai, India). The plate was inspected for growth for up to five days. Identification All toxigenic isolates were identified by conventional morphological and biochemical tests as described by Jousimies-Somer et al . [6] Sugar fermentations were carried out in anaerobic basal medium (Yeast extract - 1g; K 2 HPO 4 - 0.3g; NH 4 Cl - 1g; NaCl - 0.6g; KH 2 PO 4 - 0.3g; CaCl 2 - 0.1g; MgCl 2 - 0.2g; KCl - 0.1g; Trace element solution - 1.5 ml, Resazurin - 1mg). Morphological studies using scanning electron microscope and atomic force microscope were also conducted. Toxicity testing All the isolates were tested for toxin production by mouse bioassay as described by CDC. [7] Swiss albino mice weighing about 20 to 25g were chosen for the experiment. Using a two ml syringe fitted with a 25gauze needle, two mice were injected intraperitoneally (IP) with 0.5 ml each of the crude, non-treated culture filtrate. All mice were observed periodically for typical neurological symptoms up to three days. Ethical clearance has been obtained from the Ethical committee. PCR-based detection of bot gene Further typing of the toxin was performed by PCR amplification of chromosomal DNA of C. botulinum isolates with type specific primers. Oligonucleotide primers, selected and designed from the published DNA sequence of the BoNT gene, four new primer pairs with each being specific for C. botulinum strains were used. The primers were selected from the unique regions of each of the BoNT types A, B, and E gene and the thermal conditions was used as described by Dixit et al . [5] 16S ribosomal DNA amplification For confirmation of strain, amplification of 16S ribosomal DNA was performed with C. botulinum . The universal forward and reverse primers used were 5'- AGAGTTTGATCATGGCTCAG-3' and 5'-GTGTGACGG-GCGGTGTGTAC-3' respectively. The PCR conditions of 30 cycles were 95°C for 1 min, 54.5°C for 40 sec, 72°C for 2 min and final extension at 72°C for 5 min. A control tube replacing the template with distilled water was included with every experiment. Sequencing The PCR product of 16S rDNA amplification was purified with a PCR purification spin kit as per the manufacturer's instructions (Qiagen, GmbH, Strasse 1, 40724 Hilden, Germany). The purified PCR products were then sequenced directly by using a Taq dideoxy terminator cycle automatic sequencer (ABI/Perkin-Elmer, Norwalk, Conn.) in both directions using approximately 300 ng of template DNA per reaction mixture and five pmol of forward and reverse primers. Restricted digestion of DNA Genomic DNA prepared by the freeze thaw method [8] was digested to completion by incubation of the EcoR I restriction endonuclease enzyme for one hour, at 37ºC, with 15U, under conditions recommended by the manufacturer (Promega Corporation, 2800 Woods Hallow Road, Madison, WI, USA). The enzymic reactions were terminated by heating samples at 68ºC for 10minutes. The DNA fragments were separated by electrophoresis in a horizontal gel containing 0.5% agarose (HiMedia, laboratories, Mumbai, India). To reduce the complexity of the DNA patterns, the agarose gel was electrophoresed under conditions which preferably separated out larger DNA fragments in the range 25 to 150 kilobases. The gel was run overnight at 30 V and photographed through Alpha Gel documentation system (Alpha Innotech, San Leandro, CA). Protein pattern analysis The cell pellets were re-suspended in an equivalent volume of distilled water, placed in an ice bath and sonicated for three minutes with an ultrasonic probe emitting 45w at maximum power. The whole-cell sonicates were stored at minus 70ºC. The whole-cell sonicates and culture supernatants were analyzed by a modified method of the SDS-PAGE technique of Laemli. [9] Electrophoresis was performed with a discontinuous buffer system (Bangalore Genei, India). The gel was stained in 0.25% Coomassie Brilliant Blue-R and captured through Alpha Gel documentation system (Alpha Innotech, San Leandro, CA) and stored in computer. Numerical analysis and computation of similarity The size of each DNA-amplified fragment was determined using the molecular-weight DNA marker. A similarity matrix for all pair-wise combinations of REA profiles and protein profiles were constructed using the simple matching coefficient [10] as a measure of proximity. The coefficient of Dice was calculated and derived by the formula: Average percentage similarity (%S) = {Number of matching bands x 2/Total number of bands in both strains} x 100 Then, a dendrogram was constructed from the similarity matrix by using the Neighbor-Joining method using Mega 3.1 software program. [11] Results In the present study, 115 soil samples were cultured for isolation of Clostridium. Soil samples showed a prevalence of 51.30% (59 of 115 samples) Clostridium species. Among the 59 strains obtained, 12 toxigenic strains were documented up to species level which includes C. chauvoei , C. ramosum , C. tetani , and C. novyi . The percentage of isolation of C. ramosum and C. tetani being predominant were 3.48% and 2.61% respectively. No pathogenic strains of clostridia were obtained from other soil samples. The pH of the soils ranged from 4.0 - 8.7, with a median of 6.35. There was no significant difference in the recovery of Clostridial strains in different pH of the soil samples collected from various places. From the aquatic systems, a total of 59 samples were collected from various places of slaughterhouses, harbor, industries, lakes and water bodies and 71 Clostridial strains were isolated. Among the 71 strains isolated, three toxigenic strains were documented up to species level which includes C. bifermentans and C. chauvoei were obtained respectively from Cochin harbor and slaughterhouse sewage sample. The percentage of isolation was 3.39% for C. bifermentans and 1.69% for C. chauvoei . No other pathogenic clostridia species were recovered from the lake and slaughterhouses waste water. And in the pork, beef, poultry and goat meat samples obtained from various meat processing areas such as slaughterhouses, meat markets and meat industries, four main clostridial species such as C. chauvoei (2.86%), C. botulinum (2.86%), C. novyi (1.43%), and C. bifermentans (1.43%) were obtained. Among the 140 samples, 102 clostridial strains were isolated and among them, 10 toxigenic strains were identified up to species level. Area distribution of isolates was variable as shown in [Table - 1]. The bio-chemical profiles of all clostridial strains were tabulated in [Table - 2]. The C. botulinum isolates demonstrated that they belong to non-proteolytic (Group II) C. botulinum . The toxinotyping of all the four isolates gave amplification at 727 bp indicating positive for C. botulinum type B (data not shown). The highest sequence similarity value (98%) was obtained with the 16S rDNA sequence of the C. botulinum type B strain [Figure - 1] and lower percentage of identity (97%) was observed with the other C. botulinum strains. The sequence data have been submitted to the GenBank nucleotide sequence database under the accession number: EF 602027 for 16S rrs sequence of C. botulinum . The scanning electron microscope used in this study had a magnification range of 20 to 100,000, but for bacterial spores the most useful range is from 1,000 to 50,000. At the lower magnifications, quantitative estimates of spore populations can be made, whereas detailed surface structure can be seen at higher magnifications (greater than 10,000). The average spore size was calculated as 1.14 by 0.62 ìm and the cell size was estimated as 2.5 ìm in length and 0.7 ìm in width. The results revealed by the atomic force microscopy on the size of the vegetative cell and the spore were similar to that obtained from scanning electron microscope [Figure - 2]. REA profiles produced by the isolates had uniform digestion pattern with sizes ranging from 35 to 160Kb [Figure - 3]. From the restriction patterns, a similarity matrix was generated using Dice index similarity coefficient and the dendrogram derived from the restriction pattern of the isolated yielded two clusters [Figure - 4]. In the first cluster, most of the isolates were grouped separately in two sub-clusters, and were not distinct. Most of the C. chauvoei isolates were grouped under second cluster with few strains of C. tetani , C. bifermentans and C. novyi . The patterns were reproducible in different runs at different times. To type our strains, a simple and straightforward method, originally described by Poxton et al ., [12] i.e., the electrophoretic patterns of whole-cell and excretory proteins from strains grown under standardized conditions, was used. By this method, a great variety of patterns was obtained from the strains examined. One-dimensional SDS-PAGE of whole cell protein extracts of the 27 strains studied produced patterns containing about 40 discrete bands with molecular masses of 30 to 200 KDa [Figure - 5]. From the protein patterns obtained, a similarity matrix was generated using Dice index similarity coefficient and the dendrogram derived from the distance values yielded two electrophoretic clusters in the dendrogram related to the protein profiles of the strains [Figure - 6]. The first cluster was divided into sub-clusters and strains of C. chauvoei , C. ramosum and C. novyi grouped within a separate cluster. Strains of C. botulinum , C. bifermentans and C. novyi were grouped within the second cluster. The whole-cell protein patterns of the C. novyi , C. bifermentans and C. tetani isolates were fairly homogenous. Isolates of C. ramosum , C. botulinum and C. chauvoei were also homogenous with some variability, primarily localized in the low molecular mass region (estimated molecular mass, ~30-75 KDa). The dendrogram of the whole-cell protein profiles revealed well defined clusters among C. chauvoei , C. ramosum and C. novyi isolated from different control points from different locations. Two strains of C. botulinum obtained from fish meat dump (Clbotul1) and raw fish from Cochin Harbor (Clbotul4), formed unique pattern. The results indicate the spread of genetically identical and non-identical strains of the species in the location. However, the protein profiles of C. botulinum were quite different from C. bifermentans strains and were heterogeneous. Isolates of C. tetani (Cltetan2, and Cltetan3) was placed in a group of its own due to its slightly different pattern from the other C. chauvoei . The patterns served both to group strains when common bands were evident and to differentiate these groups on the basis of pattern heterogeneity. The excretory protein patterns in contrast, were quite different from the clusters obtained from whole-cell protein profile [Figure - 7]. The similarity matrix was generated using Dice index similarity coefficient and the dendrogram [Figure - 8] yielded two clusters with all the C. chauvoei isolates grouped under a single cluster and the remaining strains grouped within the groups of first cluster. The protein patterns of the isolates examined were highly reproducible both within and between gels. Replicate protein samples and molecular weight protein standards, run on each gel, gave similarity values of 94.3% plus/minus 2.1% and 95.0 plus/minus 2.4% respectively. Discussion In the present study, we analyzed the diversity of 27 Clostridial strains isolated from soil, water, meat and associated polluted sites. Identification was performed by conventional morphological and biochemical tests and amplification of toxin gene for the toxinotyping of C. botulinum strains. In this study, high levels of Clostridial strains were recorded in meat samples and mainly during slaughter events. Municipal sewage and storm water runoff become the conduits for the passage of pathogens into surface waters. [13] Thus, municipal sewage and waste water discharges from slaughterhouses as well as poultry and pig farms, cattle shed and fish markets were included in our study. The organisms were isolated from soil, aquatic sediments and meat samples and the percentage per total in each location was different among the locations. Clostridium chauvoei was isolated from all sampling locations, while the other species were not. And were the most frequent group comprising of majority of isolated species (2.23%) from the total samples analyzed. This is a result of the species being ubiquitous in nature. [14] Previous studies [15] reported that soil played a part in determining the survival of C. chauvoei in areas of high prevalence in Southern Africa. The higher percentage isolation of C. chauvoei from soil compared to aquatic samples and meat samples can be justified from this. The presence of culturable anaerobic species such as C. bifermentans in marine sediments collected in the coastal area near the river discharge suggests that this species can survive for a long time in the natural environment. There are several botulism hazards associated with the marine environment, sea food and fish. Nearly all of the fish-borne botulism outbreaks recorded in Canada, USA, USSR, Europe, Japan and Iran have been linked to the consumption of smoked, salt-dried, canned or fermented fish usually eaten without further cooking. [16] C. botulinum type E is the most prevalent serotype among them. Seafood samples from local retail outlets in and around Cochin have been investigated earlier and found to be contaminated with 19% C. botulinum . Non-proteolytic C. botulinum type B was expected in the fish samples but none obtained in previous studies. In our study, samples from soil and other smaller water bodies along with the Cochin harbor samples were included. The isolate obtained from fish samples from Cochin harbor as well as two isolates obtained from fish market areas were non-proteolytic C. botulinum type B. For the isolation of C. botulinum strains, C. botulinum isolation agar (HiMedia, laboratories, Mumbai, India) was used with added supplements of egg yolk suspension and selective inhibitory agents including Trimethoprim, Sulphamethoxazole and Cycloserine. In this study, it was shown that there is a large diversity of toxigenic Clostridia species responsible for gas gangrene wound infections and neurological disorders within the natural soil, aquatic, and meat environment. Furthermore, some species were found higher during storm events, suggesting that they were also mobilized from widespread animal-rearing and slaughter activities within the slaughterhouse and animal rearing sheds, and for this reason, they may be good indicators of contamination sources. In the present study, the isolates were characterized by the REA pattern. However, REA profiles do not reveal stable genetic differences among the strains. In fact, REA profile tests of the isolates are generally a useful tool to understand the epidemiology. Application of REA on C. difficile strains as an epidemiological tool has been reported earlier. [17] REA is one of the useful methods for determining the relatedness or unrelatedness of bacterial strains. Thus, it should be used in combination with, but not alone, other epidemiological tools. Different restriction enzymes tested with the strains were BamH I, EcoR I, Hae III, Hind III, Sal I, Sma I and Xho I. Among them, EcoR I which produced highly resolved DNA restriction fragments was selected as the suitable enzyme. Other enzymes which gave insufficient banding to distinguish between strains were avoided. Spermidine (100mM) included in the reaction improved markedly the digestion of genomic DNA. All the strains found in this study were easily discriminated by REA profiles and protein patterns. An examination of the whole-cell protein type groupings shows that the pattern types clustered into sets that accurately differentiated the Clostridial species. This is congruent with the preliminary results obtained with other species. [18] Of 27 isolates tested, four species for which at least seven isolates were analyzed showed a single characteristic REA pattern and five showed multiple pattern types that were also unique to a C. chauvoei characteristic REA pattern. The identification of isolated strains by restriction digestion and protein patterns has proven to be a rapid and relatively low-cost method that allows the screening of a large number of strains. In some situations, this method was more sensitive than traditional serotyping and allowed the identification of strains with an incomplete serological formula. Furthermore, it was possible to discriminate species within a single serotype, suggesting genetic diversity and/or variability within a given serotype. Further investigations will be needed to understand the significance of this genetic heterogeneity. Whole-cell protein pattern analysis by SDS-PAGE is again useful for typing strains of Clostridium and, as in previous studies appears to offer a degree of discrimination better than or similar to that of most other typing schemes. [19] Different electrophoretic methodologies have been employed in the typing of Clostridium species by protein patterns, and it appears to be the most popular general approach. Typeability is extremely high, and in theory all strains can be typed; in practice, well over 95% of isolates can be typed to previously defined types and any new patterns can be added as new types. Discrimination varies according to technique and the method used to define types. Whole cell protein typing by SDS-PAGE can be successfully applied to the epidemiological investigation of Clostridium outbreaks. It is relatively cheap and is not technically complex, and all strains are typeable. Calculation of restriction pattern similarity coefficients and graphical display of pattern relatedness as dendrogram is also useful for interpretation, particularly for large scale studies. Although this quantitative analysis has been criticized as invalid for phylogenetic interferences, because DNA restriction fragment pattern variation is not due to independent events, it is supported by population analysis. Additional population genetic and micro-evolution studies are needed to provide a better understanding of the nature and frequency of molecular events giving rise to genomic polymorphisms exploited empirically by epidemiologists for strain typing. The identification of pathogens isolated from poultry and other meat systems are commonly employed in quality and hygienic control. Several previous investigators [20] have looked into various pathogens in poultry, beef, and pork carcasses and in slaughterhouses. In microbiological analysis associated with epidemiological investigation of outbreaks, it is often necessary to obtain a more detailed identification and characterization of the organisms involved that can be provided by conventional methods such as plasmid analysis and whole cell protein analysis. SDS-PAGE, usually combined with dendrogram derived from the numerical analysis of the whole-cell protein patterns of the strains, has been used extensively to study the differences among bacterial genera, species and strains. DNA restriction digestion analysis has also proved a useful method for differentiating isolates. [4] The purpose of this study was to investigate the protein patterns and plasmid profiles to allow characterization and differentiation of the identified bacterial isolates of poultry. Although a limited number of strains were studied, isolates from widely differing geographical sources are included and the isolates obtained within a particular region produced similar DNA profiles. Restriction endonuclease DNA analysis and protein profiles appears to be a more direct and definitive approach to differentiating organisms and should provide a useful epidemiological markers for the study of clostridium associated disease. This result underlines the importance of the protein pattern of the clostridial strain in determining the clustering. The protein pattern revealed a significant heterogeneity among strains of the same species such as C. botulinum and C. bifermentans . All the results of this study clearly show high discriminatory potential of the protein profile analysis in differentiating the clostridial strains isolated from the samples at the species level. Strains of the same species with similar or different protein patterns, or REA profiles, could be isolated from the same control points in the slaughterhouses and other places. This situation indicates the importance of preventing the spread of pathogens in meat processing and controlling microbial contamination in meat plants. Differentiating among different strains of the same species is a critical part of any epidemiological study, allowing us to detect outbreaks at all levels. Conventional identification methods still have an important role to play in routine microbiological testing. However, it is believed that the use of molecular tests for differentiation of bacteria is a useful approach for establishing control strategies and controlling any bacterial outbreak. References
Copyright 2009 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb09090f3.jpg] [mb09090f4.jpg] [mb09090f5.jpg] [mb09090f8.jpg] [mb09090f1.jpg] [mb09090f2.jpg] [mb09090f7.jpg] [mb09090t2.jpg] [mb09090f6.jpg] [mb09090t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}