 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
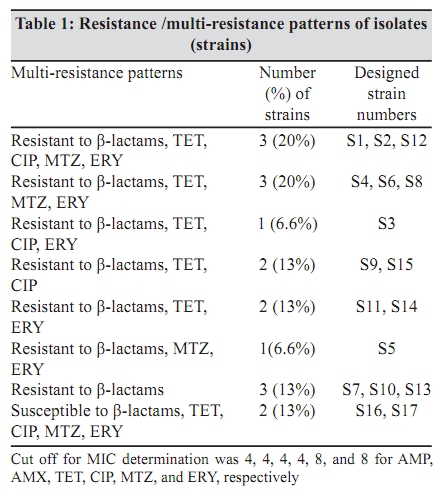
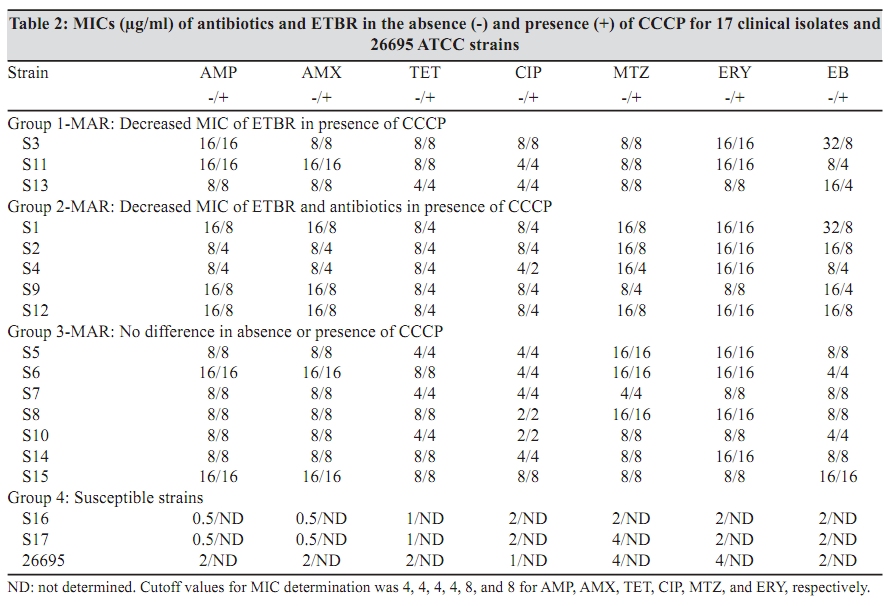
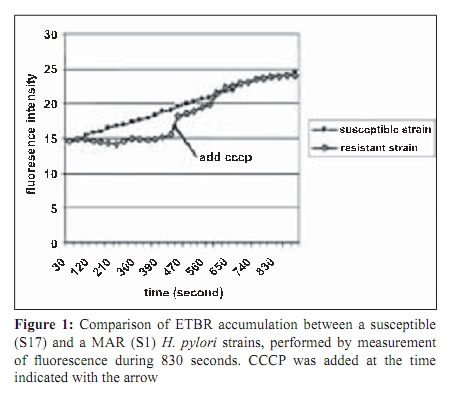
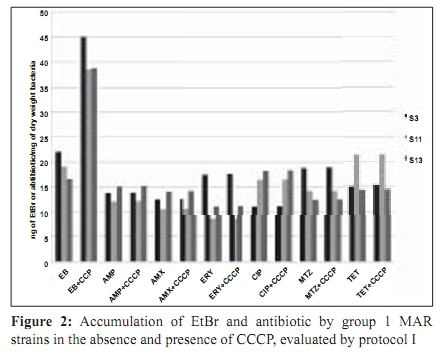
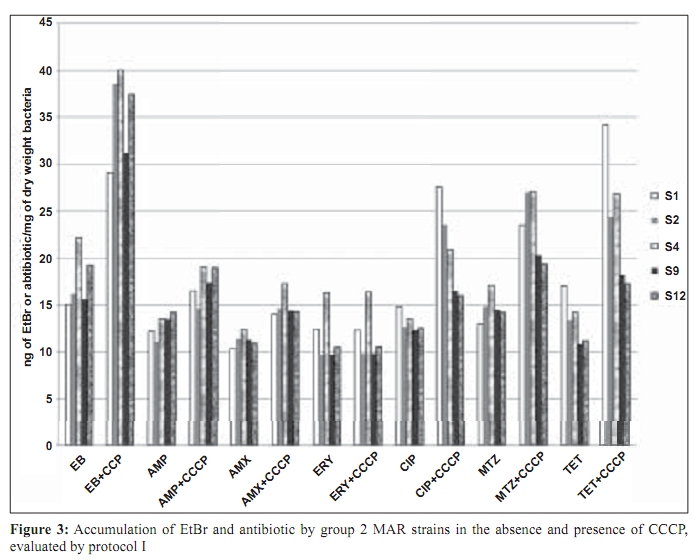
Indian Journal of Medical Microbiology, Vol. 27, No. 4, October-December, 2009, pp. 335-340 Original Article The role of active efflux in antibiotic - resistance of clinical isolates of Helicobacter pylori *T Falsafi , A Ehsani, V Niknam Department of Biology (TF, AE), Azzahra University, Tehran, Iran, Department of Biology (VN), Tehran University, Tehran, Iran Correspondence Address: *Department of Biology, Azzahra University, Tehran, Iran, tfalsafi@yahoo.com Date of Submission: 15-Jan-2009 Code Number: mb09094 PMID: 19736403 DOI: 10.4103/0255-0857.55452 Abstract Purpose: In gram-negative bacteria, active efflux pumps that excrete drugs can confer resistance to antibiotics however, in Helicobacter pylori this role is not well established. The purpose of this study is to evaluate the role of active efflux in resistance of H. pylori isolates to antibiotics.Materials and Methods: Twelve multiple antibiotic resistant (MAR) isolates resistant to at least four antibiotics, including β-lactams, metronidazole, tetracycline, erythromycin, and ciprofloxacin; three resistant to only β-lactams, and two hyper-susceptible isolates, were obtained from screening of 96 clinical isolates of H. pylori . Their minimal inhibitory concentrations (MICs) for antibiotics and ethidium-bromide (EtBr) were compared in the presence- and absence of a proton-conductor, carbonyl cyanide-m chlorophenyl-hydrazone (CCCP) using agar-dilution and disc diffusion. Drug accumulation studies for EtBr and antibiotics were assessed in the presence and absence of CCCP using spectrofluorometry. Results: MIC of EtBr for eight MAR-isolates was decreased two- to four-folds in the presence of CCCP, of which five showed reduced MICs for β-lactam, metronidazole, tetracycline, and ciprofloxacin with CCCP. Accumulation of EtBr by the MAR-isolates was rapid and not dependant on the pattern of multiple resistance. Antibiotic accumulation assay confirmed the presence of energy-dependant efflux of β-lactam, metronidazole, tetracycline, and ciprofloxacin, but no erythromycin in five MAR isolates. Energy-dependant efflux of EtBr or antibiotics was not observed for four MAR-isolates, and three isolates were resistant only to β-lactams. Conclusion: Energy-dependant efflux plays a role in the resistance of H. pylori clinical isolates to structurally unrelated antibiotics in a broadly specific multidrug efflux manner. Difference in the efflux potential of MAR isolates may be related to the presence or absence of functional efflux-pumps in diverse H. pylori isolates. Keywords: Antibiotic resistance, Active efflux, Accumulation, Helicobacter pylori, Iran Introduction Successful treatment of H. pylori infection requires two or three antibiotics, including amoxicillin (AMX), clarythromycin (CLA), metronidzole (MTZ), or tetracycline (TET), given with a proton pump inhibitor. [1] Although many H. pylori strains are quite susceptible to these antibiotics, a worldwide increase in the incidence of H. pylori isolates, resistant to a majority of these antibiotics, was observed. [1],[2] Furthermore, an increase in multiple resistances in H. pylori was observed in the isolates from children and adults that could be the major cause of treatment failure. [3],[4] Mechanisms of acquired resistance to CLA, MTZ, TET, and β-lactams, due to a (the) mutation (s) in the antibiotic target or decreased membrane permeability of the antibiotic into the bacterial cell have been studied in H. pylori . [2],[5],[6],[7],[8] However, in gram-negative bacteria, active efflux pumps that excrete drugs can also confer resistance to antibiotics. [9],[10],[11] Studies on these pumps have been done on important gram-negatives as Escherichia coli , Pseudomonas aeroginosa , Salmonella typhimurium , and Campylobacter jejuni . [12],[13],[14],[15] In the case of H. pylori , the role of the active efflux in the resistance of H. pylori isolates is not well understood. Furthermore, the limited number of works performed in this field show contradictory results on the role of active efflux in intrinsic resistance of H. pylori to antibiotics. [16],[17] The aim of this study is to evaluate the role of active efflux in the resistance of clinical H. pylori isolates to commonly used antibiotics. Materials and Methods Bacterial strains and culture conditions: Biopsies from children (median age, 9.4 years, range 5 to 15 years) admitted to Children`s Medical Centre of Tehran for upper gastrointestinal problems were used for bacterial culturing. Culture of biopsies was performed according to the previously described method. [18] Identification of isolates was performed by Gram staining, biochemical tests, and polymerase chain reaction (PCR) using the primers for H. pylori specific 16sRNA and UreC . The isolates were stored at -70°C until use. H. pylori ATCC 26695 (from Pastor Institute, France) and ATCC 43504 (from CNR, Bordeaux, France) strains were used as the control strains. Antimicrobial MIC determination: Susceptibility to ampicillin (AMP), AMX, CIP, erythromycin, (Zakaria, and Exir, Iran), MTZ, and TET (Sigma, USA) was determined by minimal inhibitory concentration (MIC) according to the clinical laboratory standards. [19] Plates were prepared with Mueller Hinton (Merck, Germany) agar plus 7% fresh sheep blood and an appropriate concentration of antibiotics from 0.5 to 64 ìg/ml. H. pylori isolates were grown for three days on modified campy blood agar plates, harvested, and suspended in sterile saline, to obtain a Mc Farland opacity of 2. The Mueller-Hinton sheep-blood (MHSB) agar plates containing antibiotics were spread with bacterial suspension and incubated for two to three days at 37°C under microaerobic conditions. Antibiotic free plates were inoculated at each series of the test to confirm the validity of the inoculums (2 ×10 8 ) and to observe for the presence of any contamination in the inoculums. H. pylori ATCC 26695 and ATCC 43504 strains were used as quality control organisms and the measurements were repeated thrice to confirm the results. The cut off values for susceptibility determination were 4 µg/ml for AMP and AMX, 8 µg/ml for MTZ, 4 µg/ml for TET, 4 µg/ml for CIP, and 8 µg/ml for ERY. Determination of the MIC was also performed in the presence of carbonyl cyanide m chlorophenyl hydrazone (Sigma, USA), an inhibitor that disrupted the proton gradient across the membrane. For this purpose, the MHSB agar plates containing antibiotics were inoculated as described earlier, dried for 5 minutes, and then blank discs containing CCCP at a concentration of 100 µM (this concentration of CCCP itself showed no collapse with bacterial growth) were placed on the agar surface. The plates were incubated for two days and at the end of the incubation period, MIC was defined as the lowest concentration of antibiotic in a plate that showed growth inhibition around the CCCP disc. EtBr MIC determination: As the agar dilution method could cause a higher risk for contamination of the environment, along with instruments by EtBr, MIC was determined by using the disc diffusion method. For this purpose, the inoculated plates (as described earlier) were dried for 5 minutes, and the blank discs containing EtBr (Merck, Germany) from 4 to 64 µg/ml were placed on the Mueller Hinton-blood agar plate. After incubation, MIC was defined as the lowest concentration of EtBr that showed a growth inhibition zone around the disc. This test was performed in a duplicated series, one for determination of MIC for EtBr without CCCP, and the other for determination of MIC for EtBr in the presence of CCCP. In case of the latter, 10 minutes after placing the discs containing EtBr, 10 µl CCCP at a concentration of 100 µM was added to the disc. After a two-day incubation of the plates, the MIC was defined as the lowest concentration of EtBr with or without CCCP showing a growth inhibition zone around the disc. Accumulation of EtBr: Accumulation of EtBr was performed using a method adopted from the previously described [14] Two-day bacterial cultures were harvested, and resuspended in prewarmed Brucella broth (37°C) to a turbidity (OD 600 ) equivalent to 0.5. After the cells were incubated for 10 minutes at 37°C, a 30-second time zero aliquot was withdrawn and EtBr was added to a final concentration of 20 µg/ml. Further aliquots were withdrawn at various intervals from 120 to 830 seconds. Accumulation of EtBr was performed in duplicated series, one without CCCP, and other in the presence of CCCP. In the latter, at 420 seconds after addition of EtBr, CCCP was added to the reaction mixture in a final concentration of 200 ìM. Fluorescence was measured with a Shimadzu RF 5000 spectrofluorometer (Shimadzu Scientific Instruments, Inc., Columbia, Md.) at an excitation of 544 nm and an emission of 590 nm. Measurement of antibiotic accumulation: Accumulation of antibiotics was measured using two different protocols, similar in inoculation and antibiotic uptake steps, but different in measurement of antibiotic accumulation: In the first one, concentration of the intracellular antibiotic was determined after extraction by 0.1 M glycine hydrochloride (pH 3.0), and in the second one, it was obtained by subtraction of extracellular concentrations from those of the initial. In both protocols, two-day cultures were harvested, and resuspended in pre-warmed Brucella broth (37°C) to an (OD 600) of 0.5. The cell suspension was incubated for 10 minutes at 37°C, and then centrifuged, washed, and resuspended in 15 mM sodium phosphate buffer pH 7.2. Antibiotic uptake assays were initiated by the addition of an antibiotic to the bacterial cell suspension at a final concentration of 10 ìg/ml. Seven minutes after antibiotic addition, CCCP was added to one-half of the reaction mixture in a final concentration of 200 µM and the other half was used as control (no CCCP). An accumulation assay for both fractions was continued until 20 minutes, and then followed using two different protocols. In the first protocol each of the collected samples were immediately diluted in 2.5 ml ice-cold PBS and centrifuged for 5 minutes at 6,000 × g at 4°C. The supernatants were discarded and the pellets were resuspended in 2 ml of 0.1 M glycine hydrochloride (pH 3.0) and stayed at 25°C for 15 to 20 hours, to extract the accumulated antibiotics. After centrifugation at 6,000 × g for 15 minutes, the supernatant was used to measure the fluorescence of the respective antibiotics and the pellet was used for measurement of the dry weight of bacteria by subtracting the initial weight of the cell-containing vial. In the second protocol (II), each of the collected samples were placed at 4ºC for 1 minute (to stop the function of the efflux pumps), centrifuged at 6,000 × g for 15 minutes, and the supernatants were used to measure the fluorescence of the respective antibiotics. As noted earlier, the pellet was used for measurement of the dry weight of bacteria by subtracting the initial weight of the cell-containing vial. The optimal innate excitation and emission wavelengths for AMP, AMX, TET, ERY, and MTZ were obtained by Shimadzu RF 5000 spectrofluorometer`s program that was 400, and 450 for AMP, AMX, and TET; 300, and 350 for ERY and MTZ. The optimal excitation and emission wavelengths of CIP were 279 and 447, respectively. In both protocols the concentrations of antibiotics in the supernatants were calculated using a standard curve of the respective antibiotics (concentrations ranging from 100 to 1000 ng) in 0.1 M glycine hydrochloride pH 3.0. The results were expressed as ng of antibiotic per mg (dry wt) of bacteria. Polymerase chain reaction: DNA was extracted using the phenol-chloroform method, as previously described. [18] PCR primers (Faza Biotech Inc., Iran) were designed on the basis of published sequences of H. pylori 16SrRNA . [4] The primers (Faza Biotech Inc., Iran), forward (5` GCTAAGAGATCAGCCTATGTCC3') and backward (5`TGGCAATCAGCGTCAGGTAATG3') were amplified using an optimized protocol to detect a 700 bps region of 16sRNA . PCR for ureC gene was performed using a commercial kit (Cinnagen, Iran) that yielded a 492 bp product according to the manufacturer`s protocol. The control for PCR was the ATCC 26695 strain. Results MIC results: On the basis of susceptibility testing 17 isolates were selected from screening of 96 isolates, of which 12 isolates (strains) showed multiple resistances to β-lactams, TET, MTZ, and / or CIP, three isolates were only resistant to β-lactams, and two isolates were hyper-susceptible [Table - 1]. In the presence of CCCP, the MICs of ETBR were reduced by two- to four-folds for eight MAR isolates (S1, S2, S3, S4, S9, S11, S12, and S13). Five out of them (S1, S2, S4, S9, and S12) showed a two-fold reduction (one dilution) in MIC for all the antibiotics except ERY. The remaining isolates including the isolates resistant to only β-lactams (S7, S10, S13), or multiple resistant (S5, S6, S8, S14) with variable resistance patterns, showed no changes in MIC for antibiotics in the presence of CCCP. According to the results of MIC determination in the absence and presence of CCCP, the isolates were classified into four groups [Table - 2] Accumulation assays: To observe the active efflux phenomenon in H. pylori , accumulation of EtBr by a MAR isolate (S1) was compared to that of a susceptible (S17) one in the absence and presence of CCCP. Presence of a proton-dependant efflux in the MAR isolate was confirmed by a rapid (30 seconds) change in the intensity of fluorescence that reached the rate of the susceptible isolate 90 seconds after addition of CCCP [Figure - 1]. Accumulation assay using protocol I, showed active efflux of EtBr for eight MAR isolates (S3, S11, S13, S1, S2, S4, S9, S12), of which five also showed active efflux for all the antibiotics except ERY. Accumulation assay by protocol II, confirmed the presence of active efflux of EtBr in the same eight isolates, and that of antibiotics in the same five isolates. The remaining isolates, including the isolates resistant to only β-lactams (S7, S10, S13), or multiple resistant (S5, S6, S8, S14), with variable resistance patterns [Table - 2], showed no efflux, either for EtBr or for antibiotics. [Figure - 2] and [Figure - 3] show the results of accumulation in ng of EtBr or antibiotic per mg of dry weight of bacteria. Discussion To observe the presence of energy-dependant efflux, MIC and accumulation of EtBr by various H. pylori isolates, resistant to multiple structurally unrelated antibiotics, was compared with those of the hyper-susceptible isolates in the presence and absence of CCCP. A decrease of MIC to EtBr associated with an increase in its accumulation was observed in the presence of CCCP for eight MAR isolates [Table - 1] and [Table - 2], [Figure - 1],[Figure - 2],[Figure - 3], suggesting that the active efflux pump (s) was (were) functional in these isolates. The results of MIC and accumulation of antibiotic showed the same association in the cases of five MAR isolates. Using two protocols for measurement of accumulation, four groups of antibiotics including β-lactams, TET, CIP, and MTZ were actively effluxed by these isolates. However the difference of antibiotic accumulation in the presence and absence of CCCP was not similar to that of MIC, which was at least two-fold. This could be related to the fact that the protocols of antibiotic measurement involved steps (centrifugation, washing of the cells, extraction, and measurement of antibiotics) that might incur some expenditure. However, three independent experiments using two protocols of measurement showed similar results, suggesting that the experiments were reproducible. Association between the phenotype of multiple-resistance and energy-dependant efflux in eight MAR isolates (groups 1 and 2: [Table - 2]) and its absence in four MAR isolates (group 3: [Table - 2]) suggests the absence of functional efflux systems in the latter group. However, the presence of various resistance patterns among them [Table - 1] indicates that the pattern of multiple-resistance did not play a role in this system. The fact that the energy-dependant efflux of antibiotics played a role in the resistance of only five out of 12 MAR isolates showed that either the multidrug efflux system was not present or functional in all of the H. pylori isolates, or it was essentially silent, unless it was under specific induction conditions. This may be the cause of the contradictory results previously obtained. In the first study, [16] the energy-dependent efflux of TET, chloramphenicol, or 1-N-phenylnaphtamine was studied, and the absence of the energy-dependent efflux in H. pylori was suggested. However, the second study [17] confirmed that H. pylori contained an active multidrug efflux mechanism. The more recent study performed in this field by Van Amsterdam et al , has confirmed the role of efflux in resistance to MTZ. [20] This study indicates that energy-dependant efflux plays a role in the resistance of clinical isolates of H. pylori to four structurally unrelated antibiotics, β-lactam, TET, MTZ, and CIP. This phenotype may not be observed in all of the H. pylori isolates and it is manifested in a broadly specific multidrug-efflux manner. Difference in efflux potential of MAR isolates may be related to the presence or absence of functional efflux-systems in diverse H. pylori isolates. Much work has yet to be done to further clarify the reasons of difference among MAR isolates in respect to their efflux behaviors. References
Copyright 2009 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb09094f3.jpg] [mb09094t2.jpg] [mb09094f1.jpg] [mb09094f2.jpg] [mb09094t1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}