 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
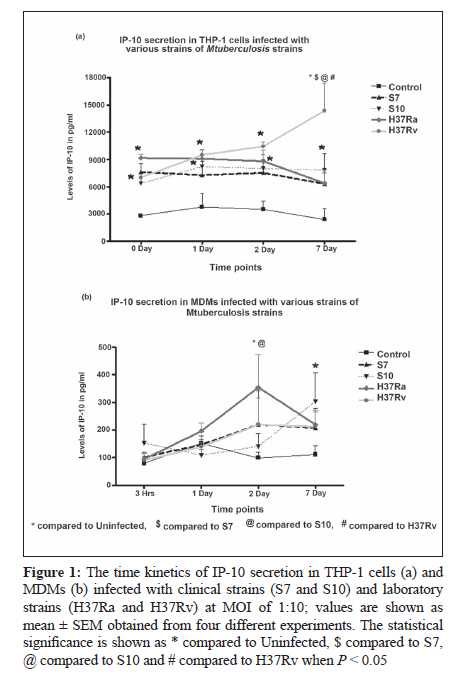
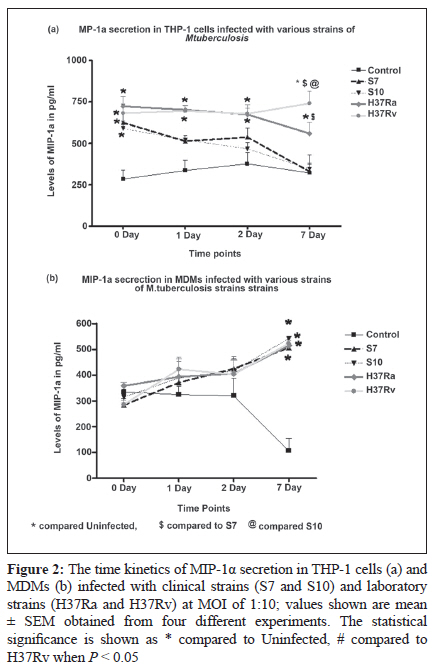
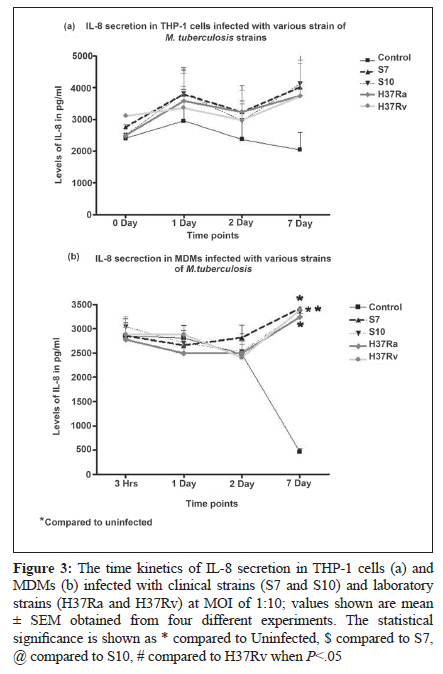
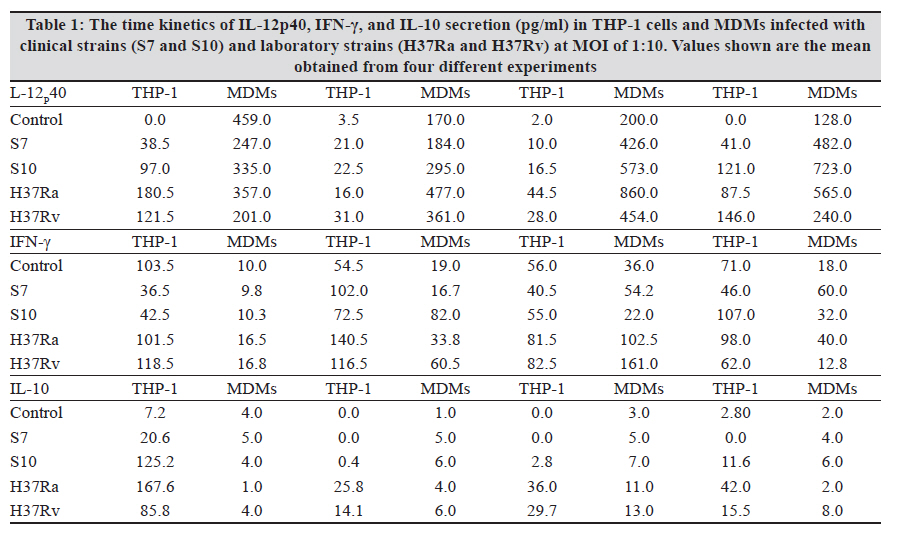
Indian Journal of Medical Microbiology, Vol. 28, No. 3, July-September, 2010, pp. 201-206 Original Article Kinetics of chemokine secretion in human macrophages infected with various strains of Mycobacterium tuberculosis P Rajavelu, SD Das Department of Immunology, Tuberculosis Research Centre (ICMR), Chennai-600031, India Materials and Methods: In this study, we used prevalent M tuberculosis clinical strains (S7 and S10) to study the chemokine secretion profile in infected THP-1 cells and monocyte-derived macrophages (MDM) and compared this with the chemokine secretion induced by laboratory strains. Results: This study showed that comparatively lower levels of IP-10 were induced by clinical strains than by laboratory strains in both differentiated THP-1 and MDMs. The secretion of MIP-1α was also depressed but only in the THP-1 cells infected with clinical strains. This depressed chemokine secretion may hinder the movement of Th-1 cells from the periphery into the infection foci to control the infection. Correlation between IP-10 and IL-12p40 showed a negative relationship in control MDMs, while there was a positive correlation in all the infected strains, indicating their cooperative role in attracting and activating Th1 cells for a protective immune response at the site. This relationship was strain dependent, with avirulent H37Ra showing higher correlation, followed by the clinical strains and the virulent H37Rv. A positive correlation of IP-10 with IFN-γ (S7 and H37Ra) and with IL-10 (H37Ra and H37Rv) suggested a definitive interplay of these molecules in infection. Conclusions: The chemokines secretion by infected THP-1 cells and MDMs was strain dependant and the lower induction by the clinical strains may indicate that the clinical strains maintain a quiescent nature to mislead the host immune system for their benefit. Keywords: Chemokine, IL-12 and IL-10, macrophage, Type -1 cytokine Introduction In tuberculosis, granuloma formation at the site of infection is one of the essential components of the host immune response for controlling the disease. Granuloma formation is a complex process and requires the activation of lymphocytes at the focus of infection and the recruitment of monocytes, naοve lymphocytes, and Th1-type cells from the blood stream into the tissues to surround the Mycobacterium tuberculosis-infected macrophages. This type of co-localization of various types of cells facilitates the activation of mycobactericidal functions in infected macrophages through the cytokines produced preferentially by the T-cells. The mechanisms for recruitment of these cells involve chemokines, which are a large family of proteins that include chemoattractants for lymphocytes, macrophages, and neutrophils. M tuberculosis infection of macrophages stimulates the production of specific chemokines for cell recruitment and activation. The alpha chemokine IP-10 is a potent T-cell attractant. [1],[2] The beta chemokine MIP-1α is an inducer of activation and proliferation of T-cells[3] and macrophages. [4] It also promotes the differentiation of Th1-type cells. [5],[6] The chemokine IL-8, which is secreted upon phagocytosis of M tuberculosis, attracts neutrophils, T-cells, and monocytes. It has been shown that the chemokine secretion upon infection with M tuberculosis is influenced by the virulence of the strain and it is suggested that the virulence-associated differences in M tuberculosis-induced chemokine secretion contributes to the failure in containing the infection due to poor granuloma formation. [7] Deficiencies in the ability to produce chemokines, especially the beta type, might lead to defects in the acquired immune response, which in turn affects the intracellular killing of M tuberculosis. [7] In addition, it has also been suggested that the expression of chemokines is probably preferentially associated with Th1- or Th2-type immune response. This is because of the differential expression of chemokine receptors, which dictates the chemokine action and their involvement in mechanisms to polarize Th1- and Th2-mediated immune responses. [8],[9],[10] In earlier studies, we demonstrated the differential ability of M tuberculosis clinical strains to induce protective immune response in healthy volunteers. [11],[12],[13] We also observed that the clinical strains (S7 and S10) maintained a uniformly spread out low-grade infection in THP-1 cells compared to laboratory strains and thereby caused no stress or strain on the infected cells. [14] This ability of intracellular pathogens to enter the mononuclear phagocytes with differential modes of phagocytosis and/or to modulate the chemokines secreted by them is probably an important mechanism for evading cellular immune responses. Hence, we aimed to study the modulation in the profile of chemokine secretion in differentiated THP-1 cells and monocyte-derived macrophages (MDMs) after infection with selected clinical strains compared to that after infection with laboratory strains. In addition, we also aimed to study how these chemokines regulated the secretion of type 1 and type 2 cytokines by these various M tuberculosis strains. Materials and Methods Bacterial strains The M tuberculosis clinical strains S7 and S10 were obtained during the Model DOTS study conducted at the BCG trial area of Tiruvallur District. These IS6110 single-copy strains were characterized epidemiologically by restriction fragment length polymorphism (RFLP) studies and were found to be prevalent in the community. [15] The laboratory strains H37Ra and H37Rv were used for comparison. Mycobacterial culture The mycobacterial strains were grown as stationary cultures in Sauton′s liquid medium until the mid-log phase of 4-6 weeks. The bacterial cells were harvested and washed and the bacterial clumps were dispersed by passing through a 26-gauge needle ten times. The cell suspension was centrifuged at low speed to remove the remaining clumps. The bacilli were adjusted to 10 Χ 10 6 cells/mL as a single-cell suspension in sterile 1x PBS and stored as aliquots at −70°C until use. The viability was checked before the start of the infection studies. THP-1 cell culture and infection THP-1 monocytic cell line was obtained from the National Centre for Cell Science (Pune, India). The cells were maintained in RPMI 1640 containing 10% heat-inactivated FCS and were differentiated into macrophages by treating with 20 nM/mL of phorbol 12-myristate 13-acetate (PMA) overnight. The cells were washed three times with plain RPMI 1640, and viable cells were seeded to a concentration of 1 Χ 10 5 in 24-well culture plates for the infection studies. The differentiated THP-1 cells were infected with S7, S10, H37Rv, and H37Ra at a MOI of 10:1 ratio for 3 hours. The cells were washed three times to remove the extracellular bacilli and the culture medium was replenished. The infected cells were incubated until the required time points (0, 1, 2, 7 days) for the assays. Uninfected cells were used as negative control. Monocyte-derived macrophage culture and infection Peripheral blood mononuclear cells (PBMC) were isolated from healthy donors′ heparinized venous blood by Ficol-Hypaque density gradient centrifugation and resuspended in RPMI 1640 supplemented with 10% autologous plasma. Monocytes were isolated from PBMC by MACS beads according to the manufacturer′s instructions (Miltenyi Biotec Inc. CA, USA). The MACS-separated population contained at least 98% monocytes as determined by flow cytometry analysis using CD14-FITC monoclonal antibodies (MoAbs). A total of 1 Χ 10 5 monocytes were seeded in 24-well culture plates and allowed to mature and differentiate into macrophages for up to 6 days for infection studies. The infection of MDMs was performed as described for differentiated THP-1 cells. Chemokine profiles The culture supernatants from the infected and uninfected THP-1 cells and MDMs were aspirated at various time points and stored at −70°C until ELISA was done. The chemokines IP-10, MIP-1α, and IL-8 were estimated by using a sandwich ELISA kit (R and D Systems, MN, USA) according to the manufacturer′s instructions. Statistical analysis The results were statistically analyzed using GraphPad Prism software (version 4.02, 2005). The data was obtained from six independent experiments, each carried out in duplicate. Chemokines and cytokines were correlated at each time point using the nonparametric correlation coefficient and compared by two-way ANOVA. Statistical significance was assumed at P < 0.05. Results In vitro chemokine secretion in THP-1 cells and MDMs after infection IP-10 The chemokine IP-10 levels in THP-1 cells after infection are represented in [Figure - 1]a. Spontaneous increase in IP-10 levels were observed in all the strains infected cells. The increase was significant in S7- and H37Ra-infected cells on day 0 in comparison to control. From day 1 up to day 7, H37Rv-infected cells showed significant increase in IP-10 levels (which was maximum on day 7) compared to the rest of the infected cells. IP-10 levels in MDMs after infection are represented in [Figure - 1]b. There was no spontaneous secretion of IP-10 upon infection up to day 1. The H37Ra-infected MDMs showed maximum IP-10 levels on day 2 and the increase was significant compared to levels in S10-infected cells and control. An interesting observation was that the clinical strain S7 and H37Rv showed almost the same levels of IP-10 at all the time points. The clinical strain S10-infected MDMs showed lower levels of IP-10 initially but the levels increased significantly on day 7 as compared to control. MIP-1α The chemokine MIP-1α levels in THP-1 cells after infection are represented in [Figure - 2]a. Spontaneous and significant increase in MIP-1α levels were observed in all the strains infected cells on day 0. The laboratory strains showed consistently higher levels of MIP-1α in THP-1 cells at all time points, whereas the clinical strains showed consistently decreased MIP-1α levels, with a significantly low level on day 7. MIP-1α levels in MDMs after infection are represented in [Figure - 2]b. There was no spontaneous increase in the MIP-1α levels upon infection on day 0. Although the infection induced the secretion of MIP-1α in MDMs on days 1 and 2, the secretion was not significantly higher than in control. Abundant secretion was seen on day 7 in all the strains infected cells and the increased levels were significantly higher than in control; however, a strain-dependent difference was not observed. IL-8 The chemokine IL-8 levels in THP-1 cells after infection are represented in [Figure - 3]a. Spontaneous secretion was observed in H37Rv- and S7-infected THP-1 cells on day 0; however, the levels were not significant. Infection with these strains indeed induced IL-8 on day 1, and the increase in secretion was maximum on day 7. A significant strain-dependent variation was not observed at any of the time points. IL-8 levels in MDMs after infection are represented in [Figure - 3]b. There was no spontaneous increase in IL-8 levels upon infection up to day 2. The maximum secretion was seen on day 7 in all the strains infected cells, and the increase was significantly higher than in control; however, a strain-dependent difference was not observed. Discussion In this in vitro study, the differentiated THP-1 cells and MDMs were infected with prevalent clinical strains of M tuberculosis to evaluate the kinetics in the secretion of chemokines as compared to the changes seen after infection with laboratory strains, namely virulent H37Rv and avirulent H37Ra. To clarify the mechanisms of establishment of protective immunity against the infection, this study also studied the influence of chemokines on type 1 cytokine secretion in both the macrophage cell types infected with various M tuberculosis strains. M tuberculosis infection of macrophages stimulates the production of specific chemokines for cell recruitment and activation. All of the beta chemokines, as well as the alpha chemokine IP-10, have been shown to recruit or activate T cells and monocytes at inflammatory foci, and all are possibly expressed following infection with M tuberculosis. Rhoades et al. studied the gene expression of alpha (MIP-2 and IP-10) and beta (MIP-1α and MCP-1) chemokine induced by different strains of M tuberculosis in in vitro and in vivo murine models. [16] They showed that the virulent Erdman strain induced lower levels of chemokines in comparison to the clinical strains CSU22 and CSU46. [16] A study by Saukkonen et al. showed that the virulent H37Rv induced significantly less MIP-1α chemokine than the avirulent H37Ra in alveolar macrophages.[7] Chan et al. have shown that human monocytes and alveolar macrophages produce IL-8 upon exposure to live M tuberculosis or the M tuberculosis heteropolysaccharide lipoarabinomannan, a putative mycobacterial virulence factor. [17] Similarly, the results from this study show moderately lower levels of IP-10 induced by clinical strains than by the laboratory strains in both differentiated THP-1 and MDMs. This depressed secretion may mean that Th-1 protective immune cells fail to migrate from the periphery into the infection loci for controlling the infection. Also, the secretion of MIP-1α was depressed only in THP-1 cells infected with the clinical strains, and neither of the infected cell types showed any strain-specific increase in IL-8 secretion. This indicates a differential pattern in the secretion of chemokines, which may be strain specific and thus influence granuloma formation and thereby hinder the protective immune response. In our previous study we showed that the THP-1 cells and MDMs infected with clinical strains induced low levels of type 1 and type 2 cytokines compared to those infected with laboratory strains [Table - 1]. In in vitro studies IP-10 induces chemotaxis of stimulated human T lymphocytes and natural killer cells. [18],[19] IP-10 is believed to be more important in the Th1-type immune response. [20] We correlated IP-10 secretion with type 1 cytokines (IFN-γ and IL-12p40) and a type 2 cytokine (IL-10) in MDMs after infection with clinical and laboratory strains to understand the influence of IP-10 in chemoattracting Th1-type cells to the focus of infection and then activating them. A negative correlation (R 2 = 0.458) was observed between IP-10 and IL-12p40 in control MDMs, while there was positive correlation in all the infected cells (S7, R 2 = 0.6995; S10, R 2 = 0.6823; H37Ra, R 2 = 0.97; and H37Rv, R 2 = 0.3643). This positive correlation clearly indicates that IP-10 (by attracting Th1 cells) and IL-12p40 (by activating them) cooperate to produce a protective immune response during infection. Although protective immunity is elicited, there is a strain-dependent response, with avirulent H37Ra showing the highest correlation, followed by the clinical strains and then by the virulent H37Rv. A positive correlation was observed between IP-10 and IFN-γ only in S7 (R2 = 0.9155) and H37Ra (R 2 = 0.9303), suggesting definitive interplay of these molecules in infection. The relationship between IP-10 and IL-10 showed a negative correlation in control (R 2 = 0.9197), no correlation in clinical strains, and a positive correlation in H37Ra (R 2 = 0.8726) and H37Rv (R 2 = 0.7779). These interesting differences in correlations between the clinical strains and the laboratory strains show how the quiescent nature of the clinical strains helps them to mislead the host immune system for their benefit. References
Copyright 2010 - Indian Journal of Medical Microbiology The following images related to this document are available:Photo images[mb10064t1.jpg] [mb10064f2.jpg] [mb10064f3.jpg] [mb10064f1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}