 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal of Biomedical Research, Vol. 3, No. 3, 2000, pp. 153 - 159 DESENSITISATION AND CALCIUM-SENSITIVITY IN THE ISOLATED PERFUSED RAT TAIL ARTERY: THE INFLUENCE OF TIME AND SOME OTHER FACTORS. ANDREW C. UGWU[1], GODFREY L. SMITH, DAVID J. MILLER AND JOHN C. MCGRATH. Autonomic
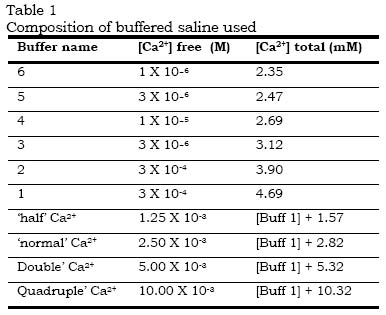
Physiology Unit, Institute of Physiology, University of Glasgow, GlasgowG12 8QQ, Scotland. Received: February 4, 2000 Code Number: md00045 The calcium sensitivity of vascular smooth muscle contraction and experimentally-induced desensitisation to noradrenaline were studied in the isolated perfused rat tail artery. Responses to the activators noradrenaline (NA) (3mM) and potassium chloride (KCl) (100mM) were obtained in Ca2+-buffered saline. Activators were added when [Ca2+] free was low (1mM) and then [Ca2+] free was increased stepwise to give a Ca2+-concentration/response curve (CRC). There was a progressive rightward shift of the CRCs with time when a series of curves was constructed. Desensitisation was not a function of time elapsed before first testing and could be reduced by increasing the interval between CRCs, reducing [NA] or doubling [glucose]. Thus the rat tail artery exhibits higher sensitivity to Ca2+ on initial contact with activators. The results suggest that desensitisation at some stage in excitation-contraction coupling, possibly by Ca2+ overload, occurs when high extracellular [Ca2+] (2.5 or 5mM) is present during activation by NA. This can be prevented by allowing recovery through leaving longer intervals between CRCs, thus allowing prolonged reproducibility of high sensitivity to Ca2+ which, otherwise, is lost. Desensitisation was essentially similar whether activation was by NA, 5HT or high [KCl]. NA caused desensitisation to 5HT more effectively than 5HT caused desensitisation to NA. The results are discussed in relation to the experimental conditions necessary to examine the sensitivity of vascular smooth muscle contraction to extracellular calcium and noradrenaline and to the mechanisms which may underline rapid desensitisation. Key Words:.Ca2+sensitivity, desensitisation, smooth muscle, noradrenaline, 5-hydroxytryptamine, rat tail arteryThe progressive fall in responsiveness of vascular smooth muscle, such as in the rat tail artery, with time or with consecutive periods of activation by agonists, has been recognized by many workers (Medgett and Langer, 1984, 1986; Spedding, 1985; Su et at., 1984). However, lack of information on the possible causes of the initial decline between the first and the subsequent responses, has led to the neglect of the first concentration - response curve; the second or even the third being taken as the control curve for the analysis of test drugs (see Su et al., 1984; Medgett and Langer, 1984, 1986; Aoki & Asano, 1986). The reason for discarding the first curve is that a significant difference usually exists between the first and the subsequent curves, thereby creating problems for analysis of the data. The basis of this difference is unclear and it is reasonable to assume that the initial state is as likely to reflect the physiological properties of the tissue as does the subsequent desensitised condition: both cannot. We have therefore made a study of the initial and subsequent sensitivity of the smooth muscle of rat tail artery with particular reference to the influence of extracellular [Ca2+] free. In addition, strategies for avoiding desensitisation are examined. A preliminary communication of some of these results has been published (McGrath, Miller & Ugwu, 1987a & b). Is it necessary to add some background on Ca pD2 values for vascular smooth muscle? METHODS Preparation of the tissue for recording perfusion pressure. 1-2cm lengths of the proximal or distal segment of the rat tail artery were prepared for recording the perfusion pressure in vitro. Male Wistar rats (300 to 350gm) were killed by a blow on the head and exsanguination. The ventral tail artery was rapidly removed and placed in aerated `calcium-unbuffered' modified krebs' bicarbonate solution. The vessel was cannulated at the proximal end and subsiquently mounted in a 5ml jacket organ bath. It was perfused with, and bathed in, saline of similar composition at 37Ec. Unbuffered saline:- The `calcium-unbuffered' saline solution was made up of the following composition (in milliomer concentration):- NaCl, 119; NaHCO3, 24.8; KH2PO4, 1.2; MgSO4.7H2O,1.2; KCL, 4.7; CaCl2, 2.5; glucose, 11.1; cocaine, 4mM; and Na2EDTA (ethylene-diaminetetra-acetic acid disodium salt), 23mM. Buffered saline:- The `calcium-buffered' saline solution had the following composition (milliomolar unless otherwise specified):- EGTA (ethelyene glycol bis-(B-aminoethyl ether) N,N,N'-tetraacetic acid), 2.5 (i.e; 0.9g.1-1); NTA (nitrilo-triaceyic acid, i.e.N,N-bis(carbonxymethyl) glycine, free acid); 2.5; NaCl, 111.5; NaOH, 7.5; NaHCO3, 24.8; KH2PO4, 1.2 MgSO4. 7H2O, 1.2; KCl, 4.7; CaCl2 was varied from 4.9 (for calcium buffer 1) to 2.35 (for calcium buffer 6); glucose11.1; cocaine, 4mM; propranolol, 1mM; and EDTA (ethylenediaminetetra-acetic acid disodium salt), 23mM. The composition of the series of buffered salines is shown in table 1. Another saline solution used in some experiments (examining the Ca2+ dependence of the contraction induced by depolarisation) had high potassium chloride, low phosphate (which allowed the use of [Ca2+]0 $ 5mM without precipitation), and had identical composition to the one described above with the following exceptions:- NaCl, 24; KCl, 100; KH2PO4, 0.1. Perfusion The preparations were tested for leakage ane those which were satisfactory were set up for perfusion. The arterial segments were mounted in the bath vertically with the cannulated proximal end of each tissue uppermost. The free distal end opened into the solution. The lumen was perfused from a gassed reservoir (370C) at a constant rate of 2-3ml/min with a pulsatile flow pump (Watson-Marlow peristaltic cassette pump, 501U with 501M multi-channel pumphead) and the perfusion pressure was recorded. This rate of flow was shown in preliminary experiments to be adequate for recording the optimal vasoconstrictor responses to noradrenaline (NA) or to high concentration of potassium chloride (KCl). The vasoconstrictor responses were measured as an increase in the peaks of the pulsatile perfusion pressure at constant average flow, using an Elcomatic EM751 pressure transducer and Devices recorder. The perfusate passing through the artery via the cannula mixed freely with the identical saline solution in the organ bath that bathed the adventitial surface. General Experimental Protocol A standard stabilisation period of 90 to 120mins in activator-free `Ca unbuffered' solution was allowed before any responses were obtained. The protocol involved changing the perfusing solution by briefly stopping perfusion and switching it to a new solution containing the required concentration of Ca2+, NA or KCl. The bathing solution in the organ bath was replaced simultaneously with the arrival of the new perfusate. Consequently both surfaces of the tissues were always exposed to an identical medium during the experiments (In practice, the respons;es to noradrenaline were no larger when it was presented to both surfaces than to the intima alone, using double cannulation (data not shown) but in order to achieve equilibrium conditions the preceding protocol was adopted). Standard exposure of the tissues to NA or KCl with either cumulative or non-cumulative protocols was for 5mins during which time the maximum response to that concentration of the activator was obtained. In some experiments, as indicated in the text, tissues were exposed to initial `priming' concentrations of noradrenaline in various buffers for 5 min before starting the main protocol. In all experiments involving responses to activators, EGTA and NTA were included (2.5mM each) so that any toxic effect would be constant throughout, and unless a particular buffer is specified total CaCl2 was adjusted to keep [Ca2+] free at 2.5mM. In all the experiments involving NA-induced contractions, cocaine (4mM) were present. Noradrenaline concentration/response curves After equilibration for 1.5 hours concentration/response curves to noradrenaline were constructed non-cumulatively, with 5min contact and 10 min wash, starting at the lowest concentration, 30nM, and proceeding in half log unit steps to 30mM. 15min intervals were left between curves in different calcium buffers, giving a routine cycle time for noradrenaline concentration/response curves of 110min. In one series of experiments the tissues were initially exposed to 3mM noradrenaline for 5 min as part of a `stabilisation' procedure, 15min before starting the first concentration/response curve. In another series this `stabilisation' procedure was omitted in order to avoid `desensitisation' (see Discussion). Calcium concentration/response curves Concentration/response curves to Ca2+ (CCRC) were constructed by changing to the lowest [Ca2+] (buffer 6; see Table 1) for 15 min, adding the activator (NA, 5HT or KCl) then, starting 5mins later, changing stepwise to higher [Ca2+] at 5min intervals starting from buffer 6 ([Ca2+]0 = 10-6 M) through a series of buffers referred to as buffers 5 to 1 which took the [Ca2+]0 to 300mM. In some experiments, further increments in [Ca2+ ] were obtained by further addition of CaCl2 to buffer 1, up to a total [Ca2+] free of 5.3mM or at times to 10.32mM. Details of the [Ca2+] in the various buffers are outlined in Table 1. The concentration of NA (3mM), 5HT (3mM) or KCl (100mM) was kept constant while changing [Ca2+] in the various buffers are outlined in Table 1. The concentration of NA (3mM), 5HT (3mM) or KCL (100mM) was kept in constant while changing [Ca2+]. At the end of the construction of the CCRC, the perfusion was stopped and the bathing and perfusion solutions were replaced by activator-free buffer 6 solution and allowed to re-equilibrate (to baseline response) before constructing another CCRC. Preliminary experiments showed that washing and "resting" buffer 6 minimised desensitisation, i.e. it was worse if [Ca2+] was higher. Intervals of 15min were allowed between curves in the initial protocol but this was varied in other experiments as noted in the next. A total time of 45minutes was taken to complete the construction of each control curve of 1 circle per hour. In one series of experiments, to test the desensitisation to a prolonged exposure to noradrenaline 3mM at a single calcium level, the time protocol was kept identical but the entire procedure was carried out in a free calcium level of 2.5mM (see figure 2). Drugs and chemicals The following substances were used:- (-)-noradrenaline bitartrate salt (Sigma), 5-hydroxytryptamine creatine sulphate (Sigma), Cocaine HCL (McCarthys), DL-Propanol HCL (I.C.I.), E.D.T.A. (B.D.H.), E.G.T.A (ethylene glycol bis-{B-aminoethyl ether} N,N,N'-tetraacetic acid) (Sigma), N.T.A.(nitrilo-triacetic acid, i.e. N,N-bis {carboxymethly} glycerine, free acid) (Sigma). Stock solutions of drugs were dissolved in distilled water, v/v and diluted in the appropriate saline. Expression of data All results have been expressed, or represented on graphs, as the mean " S.E.M. Statistical analysis was performed using Student's `t' test for paired or unpaired data, as appropriate and the 0.05 level of probability was regarded as significant. Before sensitisation occured the preparation characteristically showed a `peaked' concentration/response curve to agonists, or to calcium concentration at a fixed concentration of agonist. Both `sensitivity' and maxima subsequently declined, if steps were not taken to avoid desensitisation and, in many cases, a `maximum' was not obtained within the possible range of soluble calcium. This raises problems in quantifying calcium sensitivity, since there was often no maximum, within a given curve, on which to base the 50% response for calculation of a pD2 value. Furthermore, after desensitisation, the curves sometimes did not attain 50% of the maximum from the first curve. For simplicity we have expressed sensitivity in each curve in relation to the calcium concentration which allows a response of 30% of the maximum obtained in the first curve constructed, i.e `EC30' = concentration of calcium allowing a response of 30% of initial maximum, obtained by graphical interpolation. This corrects for inter-tissue variations in the absolute size of responses but makes no assumptions about the history of the preparation after the first curve. Thus as desensitisation proceeds, the EC30 steadily increases and -log EC30 falls. For each group of experiments sensitivity is expressed as the arithmetic mean of the -log EC30 values s.e.mean (see figues 3-6). RESULTS Responses to noradrenaline in kreb' bicarbonate saline Noradrenaline, in `normal' free calcium for kerbs' bicarbonate saline (free 2.5mM), cocoine (4mM), propanolol (1mM), EDTA (23mM), NTA (2.5mM), produced concentration-dependent contractions resulting in a sigmoid shaped concentration-response curve with a fall-off in response at high concentrations. Subsequent curves, repeated with 15-minute gaps showed a decline both in maximum and sensitivity (fig 1a). If the preparations were pre-exposed to 3mM NA for 5min in order to "sensitise", "stabilise" or "prime" the tissue before construting the concerntration respose curve to NA, the responses to successive curves were more reproducible. When three such priming exposures were carried out before construting the NA CRCs, the responses become highly productible but tended to be smaller in magnitude (fig. 1b). The later sensitivity and reproducibility of NA-induced responses were similar to those reported by Medgett and Langer (1984 & 1986). Ca2+ sensitivity of contractile responses to NA and K+ Arteries were exposed to concentration of 3mM NA (fig.2a), 100mM K+ (fig 2b), or 3mM 5HT (fig2a) in the presence of low concentration ions in solution (buffer 6). Cumulative increases of [Ca2+] free up to 5.32mM (twice the calcium concentration commonly employed), elicited concentration-dependent contractions. Six such Ca2+ concentration response curves (CCRC) were obtained with 15 minute intervals (1 hour cycle). In this series, the sensitivity of the tissues to [Ca2+] free steadily declined. With NA, the -log EC30 values were 4.4" 0.1 in the curve, declining to around 3.0, e.g 2.9 " 0.1 in the sixth curve. With KCl the corresponding values were 3, 9, " 0.1, respectively (n = 8) (see figures 2 a & b). Thus the initial but not the subsequent sensitivity to Ca2+ was greatly with NA than with KCl. A similar trend was found with 3mM 5-HT as for 3mM NA, with sensitivity declining with subsequent curves (Fig. 2c). Influence of initial incubation period An initial stabilisation period of 5 hours allowed the same sensitivity as the usual 1.5 hour period (Fig.3a). This indicates that high sensitivity remains if the tissue is not activated. Short intervals between subsequent CCRCs produced similar desensitisation after 5 hours as after 1.5 hours. Thus the deterioration in sensitivity is a function of activation and not a general deterioration with time. Influence of length of interval between CCRCs. In the experiments described above, the protocol allowed only a short rest period of 15min between exposures to NA. Increasing the gap between CCRCs reduced desensitisation still occured, though more slowly (fig. 4a). Increasing the interval by 2 hours after 3 CCRCs at 15min gaps partially arrested deterioration and allowed some restoration of sensitivity after a 2nd such interval (Fig. 4b). This suggested that 2 hr is insufficient to allow full recovery to initial sensitivity, but indicated that some recovery towards initial levels is possible. Virtually no desensitisation occured when 3.5 hours was allowed between the 1st and 2nd CCRCs: it did occur when a subsequent short gap of tissue recovers its original high sensitivity after 3.5 hours. Effect of Activation in Low [NA] When the tissue was activated with a lower [NA] (0.3mM) rather than the 3mM regularly used in this study for constructing CCRCs, desensitisation still occured but this was significantly less than that with 3mM NA (Fig. 5). Effect of "Double" [Glucose] When the concentration of glucose in the standard saline (11.1mM) was doubled and CCRCs constructed under 3mM NA activation, desensitisation was significantly reduced when compared with the controls, giving levels of calcium-sensitivity nearer to the CCRCs where normal [glucose] was used but the tissue was activated with 0.3mM NA (Fig.5). Effect of Activation by 5-HT Desensitisation of the calcium sensitivity with 5-HT (3mM) was similar to that with NA (3mM). However, with 5-HT, initial calcium sensitivity was significantly higher at very low calcium levels than with NA (Fig.2c). Their sensitivity were, however, similar in subsequent curves. Cross-desensitisation was examined. 5-HT (3mM) tested after three consecutive NA CCRCs, was desensitised. However, NA (3mM) given after three consecutive 5HT CCRCs, did not show desensitisation (Fig.6) but that recovery occurred with time. Our initial protocol, with only 15min gap between the end of one CCRC and the start of the next, allowed insufficient recovery so that desensitisation became cumulative. However, although 1 hr intervals reduced the rate of desensitisation, it was necessary to allow a very long gap of 3.5 hrs for complete recovery. It seems unlikely that such gaps would be employed by most workers so we assume that this type of desentitisation will commonly be present. Long intervals between CCRCs attenuated desensitisation and also allowed some recovery of already desensitised tissues. It has been noted in guinea-pig intestinal smooth muscle that the degree of desensitisation and the time needed for recovery are directly related to concentration of the activator (substance P) and contact time (Holzer and Petsche, 1983). Similarly, in the current study, reducing [NA] reduced desensitisation. Thus the degree of desensitisation is directly related to the strength of the activation. The partial arrest of desensitisation by doubling the glucose concentration in the saline suggests that desensitisation is partly a function of metabolic processes. This may be analogous to the "calcium overload" responsible for deterioration, in the course of experiments, of the contractile responses in cardiac muscle responses following Ca2+-overload could be linked to the inability of the cell to continuously cope with the maintenance of Ca ion efflux at the same rate as influx. Usually increased Ca2+ influx into the cell would be expected to cause increased responses, within a limit, but this influx must ultimately be balanced by rate of efflux of the ions from the cell. Initially, during the first CCRC, the cells can maintain this equilibrium. Even as this first curve proceeds, particularly into high [Ca2+] levels, the increase in response alters, as "overload" occurs. By the subsequent curves, "fatigue" has become evident as the cell strives to restore equilibrium, having partially depleted its energy reserve. At this point the increase in the cell's substrate needs in the form of glucose restored part of this lost energy and the cell was therefore able to cope with the Ca ion influx/efflux system. Consequently, desensitisation was partially arrested. The one-way cross-desensitisation between NA and 5HT was interesting. The desensitisation per se to 5HT showed that desensitisation happens with other receptors and the desensitisation to KCl showed that the phenomenon of desensitisation was not exclusive to receptor-mediated events. However the ability of NA to desensitise to 5HT together with the inability of 5HT-induced desensitisation to desensitise to NA suggests that whatever is the critical locus in desensitisation, 5HT acts on only a sub-set of that acted upon by NA. This might arise, for example if 5HT receptors were associated with a sub-population of the calcium channels which can be activated by NA. In conclusion, we have established several factors which influence desensitisation in the rat tail artery. Our results are consistent with a desensitisation in the rat tail artery. Our results are consistent with a desensitisation of Ca2+ channels by Ca2+ overload when high Ca2+ (2.5 or 5mM) is present during, or subsequent to, activation by NA thus causing reduced sensitivity in subsequent responses. This can be causing reduced sensitivity, it does seem that the initial high sensitivity to Ca2+ is a state which is "normal" for this tissue in vitro unless it is exposed to the vigorous insult of a prolonged contraction by a non-physiological concentration of NA at a high [Ca2+]. REFERENCES
© 2000 - Ibadan Biomedical Communications Group |

{kind=link}