 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal of Biomedical Research, Vol. 9, Vol. 1, 2006, pp. 57-62 Full Length Research Article Bioautographic Evaluation of Antibacterial Metabolite Production by Wild Mushrooms *Shittu, O.B1. Alofe, F.V.2 Onawunmi, G.O.3 Ogundaini, A.O.4 and Tiwalade, T.A.4 *1Department of Microbiology, University of Agriculture,
Abeokuta; 2Department of Microbiology, Obafemi Awolowo University,
Ile-Ife; 3Dept. of Pharm. Microbiology, Obafemi Awolowo University,
Ile-Ife, Nigeria; 4Dept. of Pharmaceutical Chemistry, Obafemi
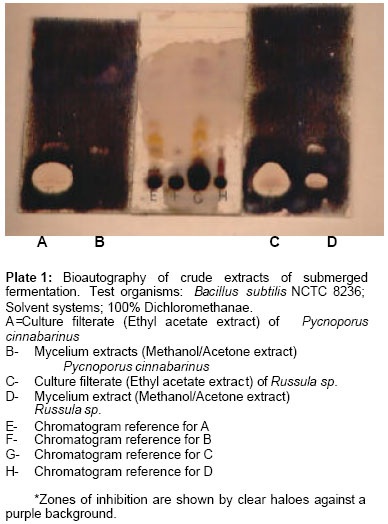
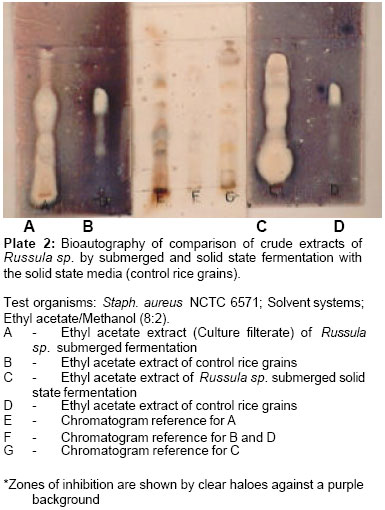
Awolowo University, Ile-Ife, Nigeria. Received: February, 2005 Code Number: md06009 ABSTRACT Russula sp. and Pycnoporus cinnabarinus were subjected to liquid and solid state fermentation for metabolite production. Local rice substrate was used for the solid state fermentation to provide a cheap and readily available medium for laboratory cultivation of wild mushroom. Bioautography, a technique that combines chromatography with bioassay in situ allows the localization of the active constituent. The result showed that in submerged fermentation, both the culture filterate and mycelial extract of Russula sp. displayed antimicrobial activity while only the culture filtrate demonstrated activity in Pycnoporus cinnabarinus.. The activity of the solid state fermentation was exhibited by the ethyl acetate fractions while the aqueous fraction showed no activity. The comparison of the culture filtrates (ethyl acetate fractions) of submerged fermentation and the ethyl acetate fractions of solid state fermentation displayed similar activities. Keywords: Bioautography, Mushroom, Metabolite, Solid state, Fermentation. INTRODUCTION Basidiomycetes produce a series of biologically active compounds when grown in pure culture (Arnone et. al., 1997). They provide a rich and varied source of terpenoids that have antibiotic and antifungal properties (Ayer and Browne, 1981). A review of literature on metabolites from mushrooms by the same workers revealed antibiotic properties of mushrooms such as Stereum complicatum, Coriolus consors, Clitocybe illudens (the jack-o-lantern mushroom), Marasmus conigenus, Lactarius rufus, Russula sardonia, Cyathus helenae, Cyathus earlei, Pleurotus mutilis, Pleurotus passeckerianus and Phlebia strigosozonakal. Solid cultures of Clitocybe hydrogramma produced marasmane sesquiterpenes that possessed antibacterial (Arnone, et al., 1997; Nasini and Vajna de Pava, 1997) and antitumor (Nasini and Vajna de Pava, 1997) activities. A compound named Oosponol, a prominent toxin with antibiotic activity, had been isolated from Oospora adsringens, Lenzites subferoginea, L. trabea, L. thermophila, Gloeophyllum sepiarium and G. abietinum (Soonenbichler et al., 1997). This compound exhibits strong antifungal activity (Chapman and Hall, 1998). Cinnabarine, a pigment of Trametes cinnabarina synonymous to Pycnoporus cinnabarina (Laessoe, 1998), Coriolus sanguineus, Polystictus cinnabarinus and Pycnoporus sanguineus showed activity against Staphylococcus aureus and fungi (Chapman and Hall, 1999). Abate and Dagne (1995) isolated two novel sesquiterpenes antimicrobial antibiotics from the wood inhabiting mushroom, Lentinus critinis. The same workers also reported the isolation of three antifungal agents from two polypores, Trametes pubesceris and Ganoderma lucidum. Dagne et. al. (1994) had earlier described the isolated antifungal compound from the culture filterate of the fungus Trametes pubescens as a polyacetylenic compound. This work was carried out to evaluate the production of antibacterial compound by wild mushrooms cultivated by submerged and solid state fermentation on local rice media using bioautographic assay. Bioautography is a technique that combines chromatography with bioassay in situ. It allows the localisation of the active constituent. The aqueous solution of a dehydrogenase indicator (2.5mg/ml-thialzolyl blue (methyl thiazoyl tetrazolium chloride) used in spraying the bioautography plates aids in visualisation showing clear haloes surrounded by blue background. The tetrazolium salt is cleaved by dehydrogenases to yield a purple formazan product thus indicating zones of growth inhibition of the test organism; the diameter of the halo increases in proportion to growth inhibition (Bengue and Kline, 1972; Rohalison, et al., 1991; Fish and Codd, 1994). The use of the tetrazolium salt in this study as a dehydrogenase indicator for detecting low metabolites down to 0.1% of the solids in the supernatant had been reported by Fish and Codd (1994). MATERIALS AND METHODS Submerged Fermentation (Cultivation On Conventional Liquid Media) Mycelial culturing, extraction and isolation of the crude extracts A method described by Anke et al. (1977) was used. A small square of 2 x 2mm of the sterile mycelial culture of each of the mushrooms was maintained on Sabouraud dextrose agar slant was used to inoculate each 250ml conical flask containing 150ml of Sabouraud dextrose broth. The flasks were incubated for 4 days on a rotatory shaker at ambient temperature and 120revs/min. The 4-day old culture were collected and filtered in rotatory vacuum filter using Gallenkamp sintered glass no.3 Buchner funnel. The mycelial pellets were washed with distilled water trice to remove traces of broth and then extracted with methanol:acetone (1:2). The culture filtrates were extracted with equal volume of ethyl acetate by partitioning in a separating funnel (solvent-solvent extraction). Both the culture filtrates (ethyl acetate extract) and the mycelial extracts (methanol:acetone extract) were separately evaporated to dryness in vacuo. Antibacterial test of the crude extracts The dried extracts obtained from above were reconstituted in minimal amount of 50% aqueous methanol and were then tested using bioautography method. The aqueous fractions were pooled and tested for activity using agar diffusion method to determine the solubility of the bioactive compounds in the organic solvent and the aqueous medium. The yields of the two ethyl acetate extracts on liquid media were recorded. Preparation of rice grains for solid state cultivation A method of preparation, described by Abate (1999) was employed. A weight of 200g of local rice grains, “Ofada”was soaked in 200ml of distilled water for three hours in 1litre conical flasks. The rice was drained and autoclaved at 121oC for 25minutes. Mycelial culturing, extraction and isolation of the crude extracts The cultivation method described by Oei (1991) and the extraction procedure described by Abate (1999) were employed. Conical flasks containing sterile rice grains were inoculated aseptically with small squares of 5x5mm mycelial culture from the full-grown pure agar culture. A control flask (un-inoculated) was also set up. The flasks were kept at ambient temperature for 14 days. The colonized rice grains and the control were separately ground with mortar and pestle and then air-dried for three hours. The ground grains were then extracted with methanol for 30minutes after which they were filtered and concentrated. The concentrates were extracted further with ethyl acetate (solvent-solvent extraction). Both the ethyl acetate extracts and the aqueous fraction were separately concentrated to dryness in-vacuo. Antibacterial test of crude extracts Thin layer chromatography studies The study was carried out to aid in the selection of the appropriate solvent system for bioautogrpahy and also to provide relevant information on the chemical properties of the mushroom species. Each extract was reconstituted in minimal amount of aqueous ethanol before spotting on the chromatography plates with silica, alumina or cellulose as the adsorbent. Each of the adsorbent was tested to select the most suitable for the bioautographic evaluation study. The plates were placed in chromatographic tanks containing solvent systems of varying polarities. The developed plates, after air-drying, were examined under the UV lamp at wavelengths of 254nm and 366nm. The plates were then sprayed with each of the appropriate chromogenic agents (Draggendorf reagent, ferric chloride, vanillin/sulphuric acid and ninhydrin). The plates sprayed with vanillin/sulphuric acid and ninhydrin were heated at 100oC until colour developed. Bioautography studies The bioautography procedure of Rohalison et al. (1991) was employed. Chromatography plates used were pre-coated with silica gel (0.2mm Kieselgel 60 F254, Merck) and glass-backed. The samples were loaded on the plates in bands. They were developed in duplicates in selected solvent systems for each of the mycelial extracts. After air drying overnight, each of the plates was placed in a humid chamber and overlaid with 10ml molten nutrient agar seeded with 0.2ml of each of Bacillus subtilis NCTC 8236 and Staphylococcus aureus NCTC 6571. Adequate humidity was maintained by placing moist cotton buds at the corners of the plates. The overlaid plates were left for 30 minutes after which they were incubated at 37oC for 24 hours. The cultures were sprayed with an aqueous solution of a dehydrogenase indicator-2.5mg/ml thialzolyl blue (methyl thiazoyl tetrazolium chloride). The plates were further incubated at 37oC for 4 hours and thereafter sprayed with absolute ethanol to kill the test organisms in order to conserve the bioautography plates. The plates were allowed to air dry and covered with plastic plates. A reference plate (not overlaid with the culture medium) sprayed with vanillin/sulphuric acid was compared with each of the bioautography plates to ascertain the location of the active constituents of the extracts. RESULT At the end of the 14-day incubation period, the grain substrates were completely ramified by the mycelial strands of Russula sp. and Pycnoporus cinnabarinus. The mycelial growth appeared within 24hours of inoculation for both fungi with distinct threads radiating from the inocula. The white mycelia covered the entire surface of the substrates as well as penetrating deep, reaching the bottom of the flask within eight to ten days. Russula sp. showed a faster growth with the fine, fluffy white mycelia becoming dull as growth advances and turning grayish after completing its ramification. Pycnoporus cinnabarinus on the other hand showed cinnabar-red pigmentation around the inoculum as well as brown coloration in the older mycelial section as growth advances. The yields of the ethyl acetate extracts by submerged fermentation for Russula sp. and Pycnoporus cinnabarinus were 19 and 23mg/ml while those of solid state fermentation were 0.719 and 0.971g/500g respectively. The yields of the crude extract were greater in the solid state fermentation. The bioautography results obtained using Bacillus subtilis as the test organism for both the mycelial extract (methanol/acetone fractions) and the culture filtrate extract (ethyl-acetate fractions) of Russula sp. and Pycnoporus cinnabarinus studied are shown in Plate 1. The clear zone of inhibition is shown against a purple background. The result revealed that the culture filtrate extract of Pycnoporus cinnabarinus exhibited a strong activity at the origin and a very weak activity at Rf value of 0.19 (Plate 1A) while the mycelial extracts showed very weak activity at Rf value of 0.19 (Plate 1B). The culture filtrates of Russula sp. on the other hand also demonstrated a strong activity only at the origin while the mycelial extract showed two zones of inhibition, a stronger one at the origin, and a weaker one at Rf value of 0.19. The bioautographic evaluation of the ethyl acetate fractions and the aqueous fractions of the solid state extracts of Russula sp. and Pycnoporus cinnabarinus using B. subtilis as the rest organism showed that only the ethyl acetate fractions of both fungi demonstrated activity while no activity was observed from the aqueous fraction. Plate 2 shows the bioautographic evaluation of the extracts of submerged and solid state fermentation of Russula sp. with the solid state media (control rice grains) for Russula sp. using Staph. aureus as test organism. The bioautography shows similar pattern of inhibition for the extracts of Russula sp. (Plate 2A and 2C). Submerged fermentation is represented by A while solid state fermentation is represented by C with Rf values of both as 0.71 and 0.9 respectively. The control rice extract represented by B and D also exhibited activity against the test organism (Plate 2B and D) showing two zones of inhibition with Rf values of 0.35 and 0.71 respectively. Plate 3 shows the bioautographic evaluation of the extracts of submerged and solid state fermentation of Pycnoporus cinnabarinus with the solid state media (control rice grains) using B. subtilis and Staph. aureus as test organisms. The extracts of rice (B and D) showed into similar zones of inhibition in both plates with Rf values of 0.3 and 0.7 (Plate 3B and 3D). The extracts of Pycnoporus cinnabarinus on the other hand gave different patterns of activity with the two test organisms. With B. subtilis, the extracts of submerged fermentation gave a zone of inhibition at the origin (Plate 3A). The solid state extract (Plate 3C) gave three haloes of inhibition (Rf values of 0.0, 0.3 and 0.7) with Staph. aureus as the test organism. DISCUSSION From the mycelial cultures obtained above, fermentation by two techniques was employed as a source of culture extracts for the production of antimicrobial metabolite(s) namely: submerged fermentation in a shake flask culture and solid state fermentation. The two fungi were successfully cultured by the two methods. Culture extract is usually reproducible and has the advantage of making possible subsequent re-fermentation to produce more materials for further biological testing and structural elucidation. Freshly grown culture ensures that there is no degradation of active components due to long term storage (Sanchuz-Puelles and Elson, 1997). Solid state fermentation had been used for upgrading the feed values of waste cellulosic materials, for the production of enzymes, aflatoxins, oriental foods and single-cell proteins (Cannal and Moo-Young, 1980; Chahal, et al., 1983). On the other hand solid state fermentation offers the advantage of cheap materials based on simple and locally available materials. Submerged fermentation is considered to be economically expensive. The results obtained by the two methods agreed with the previous reports on higher yields by solid state fermentation against lower yields by submerged fermentation (Lambert and Meers, 1983; Losane and Ramesh, 1990; Abate, 1999). However, purification of the crude extracts might give different relative yields of pure metabolite(s). The lower yields by submerged fermentation have been attributed to the use of water immiscible solvent such as ethyl acetate for extraction (Sanchuz-Puelles and Elson 1997; Abate, 1999). For submerged fermentation, 100% dichloromethane was the most suitable solvent system against dichloromethane/ethyl acetate (8.2) for the solid state extracts which was used in comparison of the extracts from the two methods. From the activity displayed by the extract of submerged fermentation the activity of Pycnoporus cinnabarinus resided in the ethyl acetate acetate fraction of the culture filtrate extract. For Russula sp. a zone of activity was given by the ethyl acetate fraction of the culture filtrate while two zones of activity were given by the mycelial extract thus indicating that both extracts posses antimicrobial activity. These implied that the antimicrobial metabolite is produced extracellularly in Pycnoporus cinnabarinus while it is both intracellularly and extracellularly produced in Russula sp. That fungal antimicrobial property can be exhibited by either culture filterate extract or mycelial extract or both finds support in various reports (Anke et al. 1976, 1977; Dagne et al., 1994; Goyal et al. 1988 ; Coletto and Labollita 1997; Ayer and Browne 1981; Inchausti et al. 1997). The compounds responsible for the bioactivity in the rice were shown as two spots on reference plates: one as a blue spot (Rf of 0.24, minor) and the second as purple spot (Rf of 0.84). Chapman and Hall (1999) have reported antimicrobial activity for rice. A compound named oryzalide A has been described as an antimicrobial constituent of rice particularly those that are resistant to Pyricularia oryzae infection. 11-Hydroxy-9-undercanoic acid and 12-hydroxy-9-undecanoic acid have also been reported as antifungal agents from constituents of the leaves of wild rice (Oryza officinalis). Thus, the antimicrobial activity recorded for the extract from solid state fermentation was due to both the fungi and the rice medium used. It was not however determined whether the activity was synergistic or additive. It has been claimed that cinnabarrine, the antibacterial and antifungal compound isolated from Pycnoporus cinnabarinus and related species, is a red pigment (Chapman and Hall, 1999). However, no colored spot as an evidence of pigmentation was obtained on the thin layer chromatography plates of the extract prior to spraying with detecting agents. This study has shown that the local rice substrate can be used in laboratory cultivation of wild mushrooms for studies in metabolite production and the detection of antimicrobial activity of the metabolite. The use of local rice is also cost effective because it is cheap, affordable and readily available. REFERENCES
The following images related to this document are available:Photo images[md06009f3.jpg] [md06009f2.jpg] [md06009f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}