 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal of Biomedical Research, Vol. 9, No. 3, Sept, 2006, pp. 173-180 Full Length Research Article Morphological Changes and Hypoglycemic Effects of Annona Muricata Linn. (Annonaceae) Leaf Aqueous Extract on Pancreatic Β-Cells of Streptozotocin-Treated Diabetic Rats Stephen O. Adewole1* and Ezekiel A. Caxton-Martins2 1* 2Department of Anatomy
and Cell Biology, Faculty of Basic
Medical Sciences, College of Health Sciences, Obafemi Awolowo
University,
Ile-Ife, Osun State, Nigeria Received: June
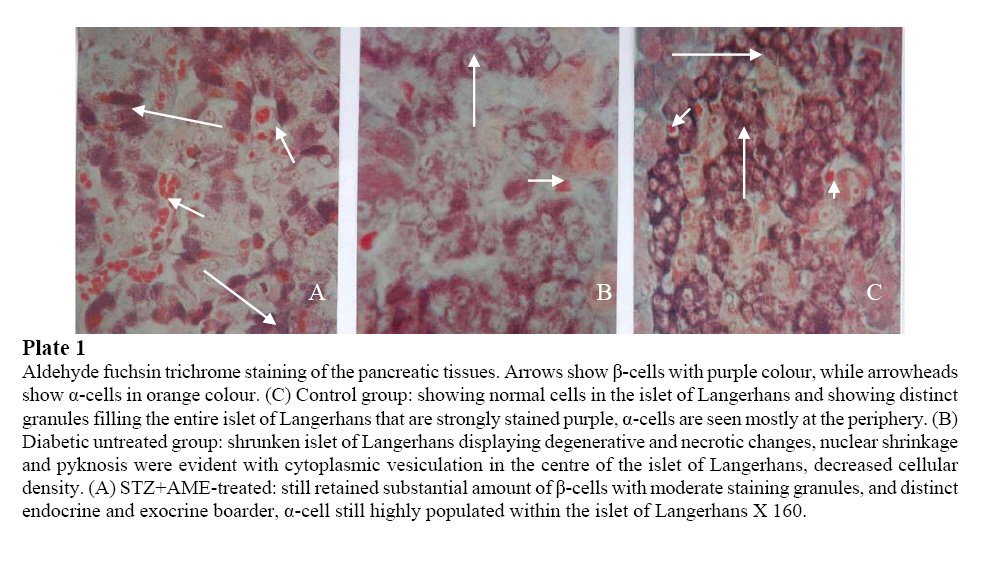
2006 Code Number: md06030 ABSTRACT The present study was undertaken to investigate the leaf aqueous extract effects of Annona muricata Linn. on the morphology of pancreatic β-cells and oxidative stress induced by streptozotocin (STZ)-diabetic rats. Diabetes mellitus was induced in the diabetic animal groups B and C by intraperitoneal injections of STZ (75 mg/kg body weight), while the control group received equal volume of citrate buffer (pH 6.3) solution intraperitoneally. The rats in group C were given A. muricata leaf aqueous extract (AME, 100 mg/kg, p.o.) as from day 5 post STZ injections, and stopped on the 30th day of the study period. The pancreases of the rats were excised and randomly processed for histological staining and biochemical assays for antioxidant enzymes [such as glutathione (GSH), superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), catalase (CAT), malondialdehyde (MDA) and serum nitric oxide (NO)]. In diabetic state, pancreatic β-cells of STZ-treated group B rats histologically demonstrated marked alterations in the micro-anatomy and cellular integrities. The morphology of A. muricata-treated rats’ pancreases showed viable cellularity with distinct β-cell mass. STZ treatment significantly decreased GSH-Px, SOD, GSH, CAT and pancreatic/serum insulin levels (p<0.05). However, STZ treatment increased blood glucose concentrations, MDA, and NO. A. muricata-treated rats showed a significant decrease (p<0.05) in elevated blood glucose, MDA and NO. Furthermore, A. muricata treatment significantly increased (p<0.05) antioxidant enzymes’ activities, as well as pancreatic/serum insulin contents. In conclusion, the findings of the present study indicate that A. muricata treatment has beneficial effects on pancreatic tissues subjected to STZ-induced oxidative stress by directly quenching lipid peroxides and indirectly enhancing production of endogenous antioxidants. Annona muricata protected and preserved pancreatic β-cell integrity. Keywords: Annona muricata Leaf Aqueous Extract; morphological Changes; Oxidative Stress; Antioxidants. INTRODUCTION Herbal remedies from medicinal plants have been used traditionally in many parts of the world where access to formal healthcare is limited. There are several reasons why the use of medicinal plants should be studied: herbal remedies may have recognizable therapeutic effects (Bailey and Day, 1989); they may also have toxic side-effect (Keen et al; 1994). Although, the use of medicinal plants provides an indication of beliefs about illness and its treatment that may conflict with beliefs of workers in the formal healthcare system (Morgan and Watkins, 1988). Over the last three decades chronic disorders such as diabetes and hypertension have emerged as the major causes of adult morbidity and mortality all over the world (Gulliford, 1994). Successful control of health problem stemming from diabetes and hypertension requires active participation of patients in their own care, but as Morgan (1995) has pointed out, health care providers’ limited understanding of patients’ concepts of illness and ideas about treatment are obstacles to establishing effective patterns of self-care. Several studies have described the medicinal purposes of Annona muricata and have outlined the social history of the plants’ use (Ayensu, 1981). A. muricata (Linn.) (family, Annonaceae) commonly called “Soursop” is a small, upright evergreen tree growing 5 to 6 meters in height. Young branchlets are rusty-hairy, the malodorous leaves, normally evergreen, are alternate, smooth, glossy, dark green on the upper surface, lighter beneath; oblong, elliptic or narrow-obovate, pointed at both ends, 6 - 20 cm long and 2 – 6 cm wide. The flowers are borne singly, may emerge anywhere on the trunk, branches or twigs. They are short stalked, 4 - 5 cm long, plump, and triangular-conical; the 3 fleshy, slightly spreading, outer petals yellow-green, with 3 close-set inner petals pale-yellow (de Feo, 1992; Vasquez, 1990). The fruit is more or less oval or heart-shaped, sometimes irregular, lopsided or curved, due to improper carper development or insect injury. The size ranges from 10-30 cm long and up to 15 cm in width and the weight may be up to 4.5-6.8 kg. The fruit is compound in and covered with reticulated, leathery-appearing but tender, inedible bitter skin from which protrude few or many stubby, or more elongated and curved soft, pliable “spines”. The tips break off easily when the fruit is fully ripe. The skin is dark-green in the immature fruit, becoming slightly yellowish-green before the mature fruit is soft to the touch. Its inner surface is cream-colored and granular and separates easily from the mass of snow-white, fibrous, juicy segments – much like flakes of raw fish – surrounding the central, soft-pithy core. In aroma, the pulp is somewhat pineapple-like, but its musky, subacid to acid flavor is unique (Schultes and Raffauf, 1990). Most of the closely-packed segments are seedless. In each fertile segment, there is a single oval, smooth, hard, black seed, 1.25-2 cm long; and a large fruit may contain from a few dozen to 200 or more seeds (Morton, 1980). It is indigenous to most of the warmest tropical areas in South and North America including Amazon, A. muricata has become naturalized in many countries, and now has a wide distribution throughout tropical and subtropical parts of the world, including western part of Nigeria. All parts of the A. muricata tree are used in natural medicine in the tropics including the bark, leaves, root and fruit-seeds. Generally the fruit and fruit juice is taken for worms and parasites, to cool fevers, to increase mother’s milk after childbirth (lactagogue), and as an astringent for diarrhea and dysentery. The crushed seeds are used as a vermifuge and anthelmintic against internal and external parasites and worms. The bark, leaves and roots are considered sedative, antispasmodic, hypoglycemic, hypotensive, smooth muscle relaxant and nervine and a tea is made for various disorders for those purposes (Holdsworth, 1990). Many bioactive compounds and phytochemicals have been found in A. muricata as scientists have been studying its properties since the 1940’s. Its many uses in natural medicine have been validated by this scientific research (Weniger et al; 1986). The earliest studies were between 1941 and 1962. Several studies by different researchers demonstrated that the leaf, bark, root, stem and seed extracts are antibacterial in vitro against numerous pathogens (Misas, 1979; Sundarrao, 1993; Heinrich 1992) and that the bark has antifungal properties (Lopez Abraham, 1979). Much of the recent research on A. muricata has been on a novel set of phytochemicals (Annonaceous acetogenins) that are found in the leaves, seeds and stem which are cytotoxic against various cancer cells (Chang, 2001; Chang, 2003; Liaw, 2002). Streptozotocin (STZ), an antibiotic produced by Streptomyces achromogenes, possesses pancreatic β-cell cytotoxic effect (Weiss, 1982). Streptozotocin has been widely used for inducing diabetes mellitus in a variety of animals. STZ causes degeneration and necrosis of pancreatic β-cells (Uchigata et al; 1982; Merzouk et al; 2000). Although the mechanism of the β-cell cytotoxic action of STZ is not fully understood, experimental evidence has demonstrated that some of its deleterious effects are attributable to induction of metabolic processes, which lead on to an increase in the generation of reactive oxygen species (ROS) (Chen et al; 1990). Apart from production of ROS, STZ also inhibits free radical scavenger-enzymes (Kröncke et al; 1995). The superoxide radical has been implicated in lipid peroxidation, DNA damage, and sulfhydryl oxidation (Tiedge et al; 1997; Matkovics et al; 1998). Oxidative stress is the excess formation and/or insufficient removal of highly reactive molecules such as reactive oxygen species (ROS) and reactive nitrogen species (RNS) (Turko et al; 2001; Maritim et al; 2003). ROS include free radicals such as superoxide (O2-), hydroxyl (OH-), peroxyl RO2), hydroperoxyl (HRO2-) as well as nonradical species such as hydrogen peroxide (H2O2), and hydrochlorous acid (HOCl) (Evans et al; 2003). RNS include free radicals like nitric oxide (NO) and nitrogen dioxide (NO2-), as well as nonradicals such as peroxynitrite (ONOO-), nitrous oxide (HNO2) and alkyl peroxynitrates (RONOO) (Liu et al; 2002). Of these reactive molecules, O2-, NO and ONOO- are the most widely studied species and play important roles in the diabetic cardiovascular complications. Oxidative stress in diabetes, sources from (i) nonenzymatic source resulting from biochemical oxidation of glucose, glucose reacts with protein in nonenzymatic manner leading to the development of advanced glycation end product (AGE) (Taniyama and Griendling, 2003). (ii) enzymatic sources which includes nitric oxide synthetase (NOS), NAD(P)H oxidase and xanthine oxidase (Guzik et al; 2002; Etoh et al; 2003). (iii) mitochondria respiratory chain, during oxidative phosphorylation process, electrons are transferred from electron carriers NADH and FADH2 to oxygen, thus generating ATP in the process (Green et al; 2004). Annona muricata has a long history of use in herbal medicine in the tropical areas in South and North America including the Amazon and West Africa. All parts of the tree are claimed to be used in natural medicine, in view of this, the present study was designed to evaluate the hypoglycemic, morphological changes of pancreatic β-cells and antioxidant effects of A. muricata leaf aqueous extract in experimental models of diabetes mellitus in rats. MATERIALS AND METHODS Animal care and monitoring: Healthy, male and female, Wistar rats (Rattus norvegicus) weighing 250–300 g (averaging 12 weeks old) and normal Mice (Mus domesticus) were used in this study. They were housed under standard laboratory conditions of light, temperature (21±2oC) and relative humidity (55±5%). The animals were given standard rat pellets and tap water ad libitum. The rats were randomly divided into three experimental groups: A (control), B (STZ-treated), C (STZ + A. muricata-treated). The control group animals (A) consisted of ten rats, while groups B and C consisted of forty rats. The mice were used for acute toxicity testing of the crude plant extract, while the rats were used for hypoglycemic and morphological evaluations of the plant’s extract. Maintenance and treatment of animals were in accordance with the principles of the “Guide for care and use of laboratory animals in research and teaching” prepared by the National Institutes of Health (NIH) publication 86-23 revised in 1985. Plant material : Fresh leaves of Annona muricata (Linn.) (family, Annonaceae) (locally known as “Soursop in English, and “Abo” in Yoruba language of Western Nigeria) were collected in Ibadan, Nigeria between April and May 2006. The leaves were identified, by the Taxonomist/Curator of the Department of Botany, Obafemi Awolowo University, Ile-Ife, Nigeria, as those of A. muricata Linn. (family, Annonaceae). Voucher specimen of the plant has been deposited in the Herbarium of the University’s Botany Department. Preparation of Annona muricata leaf aqueous extract: A. muricata fresh leaves were air-dried at room temperature. One kilogram of the air-dried leaves of the plant was milled into fine powder in a Waring blender, department of Pharmacognosy, Obafemi Awolowo University, Ile-Ife, Nigeria. The powdered leaf was macerated in distilled water and extracted twice, on each occasion with 2.5 1itre of distilled water at room temperature for 48 h. The combined aqueous extract solubles were concentrated to dryness under reduced pressure at 60oC±1oC in a rotary evaporator. The resulting aqueous extract was freeze-dried, finally giving 26.23 g (i.e., 2.62% yields) of a light green, powdery crude aqueous leaf extract of A. muricata (AME). Aliquot portions of the crude plant extract residue were weighed and dissolved in distilled water for use on each day of our experiments. Acute toxicity testing: The median lethal dose (LD50) of A. muricata leaf aqueous extract (AME) was determined in the rat using a modified method of Lorke, (1983). Mice fasted for 16 h were randomly divided into groups of eight mice each. Graded doses of AME (50, 100, 200, 400, 800, 1600 and 3200 mg/kg) were separately administered intraperitoneally (i.p.) to the mice in each of the test groups. Each of the mice in the control group was treated only with distilled water (3 ml/kg, i.p.). The mice in both the test and control groups were then allowed free access to food and water, and observed over a period of 48 h for signs of acute toxicity. The number of deaths (caused by the extract) within this period of time was noted and recorded. Log dose-response plots were constructed for the plant’s extract, from which the LD50 of the leaf aqueous extract was determined. Evaluations of hypoglycemic activity of A. muricata leaf aqueous extract (AME): The test compound (i.e., A. muricata leaf aqueous extract (AME, 50-400 mg/kg) were administered orally to the groups of fasted normal (control) and fasted diabetic ‘test’ rats. 1, 2, 3, 4 and 8 h following oral administrations of the test compound to the animals, blood glucose concentrations (Gt) were again determined and recorded. In each case and for each dose, the rats were restrained in a cage, and blood samples (0.02 ml) were collected from the tail vein of each rat for blood glucose analysis. Percentage glycemic variation was calculated as a function of time (t) by applying the formula: % glycemic change = Go – Gt x 100/Go [where Go and Gt represent glycemic values before (i.e., 0-time or 0-h glycemic values), and glycemic values at 1, 2, 3, 4 and 8 hours after, oral administrations of ‘test’ compound respectively]. Induction of experimental diabetes: Diabetes mellitus was induced (in groups B and C ‘test’ rats) by single intraperitoneal injection of STZ (75 mg/kg), freshly dissolved in 0.1mol/l citrate buffer (Rossini et al; 1978). Control rats were injected with only citrate buffer solution (pH 6.3) intraperitoneally. The ‘test’ animals in groups B and C became diabetic within 48 hours after STZ administration. Diabetic state was confirmed by measuring basal blood glucose concentration 48 hours after STZ injection. Diabetes was allowed to develop and stabilize in these STZ-treated rats over a period of 3-5 days. All animals in groups A, B and C were kept and maintained under laboratory conditions of temperature, humidity, 12-h day – 12-h night cycle; and were allowed free access to food (standard pellet diet) and water ad libitum. Before the commencement of our experiments, both the control normal (normoglycemic) and STZ-treated, diabetic (hyperglycemic) test rats were fasted for 16-h, but still allowed free access to water throughout. At the end of the 16-h fasting period – taken as 0 time (i.e., 0 h) – blood glucose levels (initial glycemia, G0) of the fasted normal and STZ-treated, diabetic rats were determined and recorded. Fasted STZ-treated rats with blood glucose concentration ≥18 mmol/L were considered to be diabetic, and used in this study. The test compound [i.e., Annona muricata leaf aqueous extract (AME, 100 mg/kg p.o.)] was administered orally (by gastric intubation) to the group C fasted diabetic rats. The administration of plant’s aqueous extract was commenced as from the 5th day post STZ injections. Histological procedures: Pancreatic tissues were excised from sacrificed animals, weighed, and fixed in aqueous Bouin’s solution for 48 h and were sequentially embedded in paraffin wax blocks according to the standard procedure, sectioned at 5 μ thickness. They were further deparaffined with xylol, and histologic observations were performed after staining for functional pancreatic tissues by Aldehyde fuchsin trichrome method described by Stevens (1982). The slides were examined using light microscope. Biochemical assays Blood Glucose and serum insulin estimations: Blood samples were obtained by repeated needle puncture of the same tail tip vein. Samples were obtained 1 day before STZ-treatment, and on various days after induction of diabetes mellitus. Diabetes was allowed to develop and stabilize in these STZ-treated rats over a period 5 days. Blood glucose concentrations were determined by means of Bayer Elite® Glucometer, and compatible blood glucose test strips (Henry, 1984). The mean fasting blood glucose levels for normal, nondiabetic rats were found to vary between 4.01±0.04 and 4.20±0.13 mmol/L. Fasted STZ–treated rats with blood glucose concentrations ≥18 mmol/L were considered to be diabetic, and used in this study. Serum insulin concentrations were determined by an enzyme-linked immunosorbent assay (ELISA) using a commercial kit (Crystal Chem, Chicago, III). Preparation of pancreas homogenates: The rats were sacrificed by cervical dislocation and pancreata were excised, rinsed in ice-cold physiological saline, and homogenized with Potter Elvehjem homogenizer. 10% homogenates were prepared in 6.7 mM phosphate buffer, pH 7.4 and centrifuged at 10,000 rpm for 10 min at 4oC, and the supernatant was used for antioxidant enzyme assays. For the determination of lipid peroxidation, pancreatic tissues were homogenized in 1.15% KCl solution to obtain a 10% (w/v) homogenate. Protein content of pancreas homogenates was determined by using bicinchoninic acid (BCA) protein assay reagent (Pierce Chemical Company, Rockford, IL). Catalase Activity (CAT): The activity of catalase (CAT) was measured using its perioxidatic function according to the method of Johansson and Borg (1988). 50 µL potassium phosphate buffer (250 mM, pH 7.0) was incubated with 50 µL methanol and 10 µL hydrogen peroxide (0.27%). The reaction was initiated by addition of 100 µL of enzyme sample with continuous shaking at room temperature (20oC). After 20 minutes, reaction was terminated by addition of 50 µL of 7.8 M potassium hydroxide. 100 µL of purpald (4-Amino-3-hydrazino-5-mercapto-1,2,4-triazole, 34.2 mM in 480 mM HCl) was immediately added, and the mixture was again incubated for 10 minutes at 20oC with continuous shaking. Potassium peroxidate (50 µL 65.2 mM) was added to obtain a coloured compound. The absorbance was read at 550 nm in a spectrophotometer. Results are expressed as micromoles of formaldehyde produced/mg protein. Reduced GSH and oxidized glutathione GSSG activities: Pancreatic GSH and GSSG contents were measured as described by Hissin and Hilf (1973). For measuring GSH contents, 4 ml of pancreatic homogenate was precipitated by adding 1 ml of 25% metaphosphoric acid and centrifuged at 10,000 x g (Ultracentrifuge, Hitachi, Japan) for 30 min. Supernatant was diluted 20 times with same buffer and 100 μl of orthopthaldehyde (OPT) was added. In addition, for GSSG assay, 0.5 ml supernatant was incubated at room temperature with 200 μl of 0.04 mol/l N-ethylmaleimide solution for 30 min and to this mixture 4.3 ml of 0.1 mol/l NaOH was added. A 100 μl sample of this mixture was taken for the measurement of GSSH using the exact procedure described above for GSH assay except that 0.1 mol/l NaOH was used as the diluent instead of phosphate buffer. Samples were incubated at room temperature for 15 min and fluorescence was measured using spectrofluorometer (Tecan Spectra Fluor Plus Germany) at 350 nm (Ex)/420 nm (Em). Superoxide Dismutase Activity (SOD): Pancreatic SOD activity was assayed by the method of Kakkar et al. (1984). Reaction mixture contained 1.2 ml of sodium pyrophosphate buffer (0.052 mM, pH 7.0), 0.1 ml of phenazine methosulphate (PMS) (186 μM), 0.3 ml of nitro blue tetrazolium (NBT) (300 μM). 0.2 ml of the supernatant obtained after centrifugation (1500 x g, 10 min followed by 10,000 x g, 15 min) of 5% pancreatic homogenate was added to reaction mixture. Enzyme reaction was initiated by adding 0.2 ml of NADH (780 μM) and stopped precisely after 1 min by adding 1 ml of glacial acetic acid. Amount of chromogen formed was measured by recording color intensity at 560 nm. Results are expressed as units/mg protein. Glutathione Peroxidase Activity (GSH-Px): Glutathione peroxidase (GSH-Px) activity was measured by NADPH oxidation, using a coupled reaction system consisting of glutathione, glutathione reductase, and cumene hydroperoxide (Tappel, 1978). 100 µL of enzyme sample was incubated for five minutes with 1.55 ml stock solution (prepared in 50 mM Tris buffer, pH 7.6 with 0.1 mM EDTA) containing 0.25 mM GSH, 0.12 mM NADPH and 1 unit glutathione reductase. The reaction was initiated by adding 50 µL of cumene hydroperoxide (1 mg/ml), and the rate of disappearance of NADPH with time was determined by monitoring absorbance at 340 nm. One unit of enzyme activity is defined as the amount of enzyme that transforms 1 µmol of NADPH to NADP per minute. Results are expressed as units/mg protein. Lipid Peroxidation contents (LPO): The product of the reaction between malondialdehyde (MDA) and thiobarbituric acid reactive substances (TBARS) were measured by a modified method of Ohkawa et al., (1979). For each sample to be assayed, four tubes were set up containing 100, 150, 200 and 250 µL of tissue homogenate, 100 µL of 8.1% SDS, 750 µL of 20% acetic acid, and 750 µL of 0.8% aqueous solution of TBA. The volume was made up to 4 ml with distilled water, mixed thoroughly and heated at 95oC for 60 minutes. After cooling, 4 ml of n-butanol was added to each tube, the contents mixed thoroughly, and then centrifuged at 3000 rpm for 10 minutes. The absorption of the clear upper (n-butanol) layer was read at 532 nm. 1, 1, 3, 3 tetraethoxy propane (97%) was used as the external standard. Results are expressed as nmoles TBARS/mg tissue. Nitric Oxide (NO) Serum nitrite/nitrate levels were determined by converting the nitrate to nitrite, using enzyme nitrate reductase followed by addition of Griess reagent to colorimetrically quantify the nitrite concentration (Green et al; 1982). The serum was diluted 1:5 in PSB before a 25-µl aliquot was added to a mixture of 25 µl nitrate reductase (1.5 U/ml) and 25 µl of NADPH (0.134 mg/ml), both prepared in 40 mM Tris, pH 7.6. The samples were thereafter incubated at room temperature for 3 hours. Following this period, 100 µl of Griess reagent (1:1 mixture of 1% sulphanilamide in 5% phosphoric acid and 0.1% naphyly-ethylenediamine) was added and incubated for a further 10 min at room temperature; the absorbency of the samples was measured at 540 nm with a 650 nm reference. The concentration of nitrite/nitrate was determined from a standard curve of sodium nitrate and calculated as µM nitrite/nitrate. Immunohistochemistry The animals were sacrificed by cervical dislocation, some at the end of the 16 hours fasting period, and others on various days following STZ administration. Pancreatic tissues were excised and weighed after the fat and lymphnodes had been removed. The splenic parts of the pancreas of each rat were fixed in aqueous Bouin’s solution, and embedded in paraffin. Each pancreatic block was serially sectioned (5 μ) throughout its length to avoid any bias due to changes in islet distribution or cell composition, and thereafter mounted on slides. For each pancreas, 10 sections were randomly chosen at a fixed interval through the block (every 30th section), a procedure that has been shown to ensure that selected sections are representative of the whole pancreas (Mossavat et al; 1997). Sections were immunostained for insulin, using a peroxidase indirect labeling technique. The sections were incubated for 1 h with guinea-pig anti-insulin serum (final dilution 1:1,000, Ref. 64-104-1; Aurora, OH). Thereafter, sections were incubated for 45 min with peroxidase-conjugated rabbit anti-guinea pig IgG (final dilution 1:20; Dako, Carpinteria, CA). The activity of the antibody-peroxidase complex was revealed with 3,3’-diaminobenzidine-tetrahydrochloride, using a peroxidase substrate kit (DAB; Biosys-Vector, Compiegne, France). A standard concentration of hematoxylin was added as a counterstain. Pancreatic insulin contents: The splenic regions of the pancreatic tissues from euthanized rats were weighed and homogenized, on various experimental days, in acid-ethanol solution (75% ethanol, 23.5% distilled water, 1.5% concentrated HCl). After overnight incubation at 4oC, the suspensions were centrifuged, and the supernatants were collected and assayed for insulin contents, using a competitive ELISA kit (Kekow et al; 1988). Plates were coated with rabbit anti-guinea-pig Ig secondary Ab (Organon Teknoka, Durhan, NC), followed by incubation with a guinea-pig anti-human insulin Ab (Cortex Biochem, San Leandro, CA). Following two washing steps, various extract dilutions or insulin standards (Linco Research, St. Louis, MO) were mixed with constant concentration of HRP-conjugated rat insulin (Organon Teknika) for 4 h at room temperature, or at 4oC overnight, before competitive capturing was allowed for 3 h. After washing five times, Sigma FAST OPD tablets (Sigma, St. Louis, MO) were used as substrate. Results were analyzed using ceres 900 C ELISA-reader. Statistical Analysis The data obtained were expressed as means (±SEM), and analyzed using repeated measures of variance. The differences between the means were analyzed statistically with one-way analysis of variance (ANOVA; 95% confidence interval). Values of p<0.05 were taken to imply statistical significance. RESULTS Characteristics of diabetic state: Forty-eight hours after STZ administration, all animals that had been treated with STZ displayed glucosurea, hyperglycemia, hypoinsulinemia and a moderate loss of body weight. Blood glucose concentrations and serum insulin levels of the STZ-treated experimental animals are shown in Table 1. The baseline weight of the rats at the beginning of the study was similar in all groups. At the end of the study period (2 months), diabetic animals in group B presented with weight loss. The initial and final body weights were not significantly (p<0.05) different in control and AME-treated groups. Blood glucose and serum insulin concentrations: Mean blood glucose concentrations and serum insulin levels of the STZ-treated experimental animals are shown in Table 1. In our control set of experiments, pretreatment of the rats with citrate buffer alone did not significantly modify (p > 0.05) the serum insulin and blood glucose concentrations. As shown in Table 2. Fig. 1, there was a gradual rise in the blood glucose concentrations of the STZ-treated rats as from day 1 following injection of STZ, and the values were significantly higher (p < 0.05) than those of control animals (Table 1). Table 1. Various parameters recorded in control, STZ-treated and STZ + AME-treated diabetic rats just before and after treatment. Values presented represent the means (± SEM) of 10 observations.
Values are expressed as means (±SEM) of 10 rats. a Insignificant difference (p>0.05) between all groups. b,c,d Significant difference (p<0.05) between treatment and control groups. Table 2. Changes in blood glucose concentrations and serum/pancreatic insulin contents in control, STZ-treated and STZ + AME-treated diabetic rats.
Values are expressed as means (±SEM) of 10 rats. a Insignificant difference (p>0.05) between all groups. b,c,d Significant difference (p<0.05) between treatment and control groups. Table 3. Pancreatic tissue CAT (μmol/mg protein), GSH (U/g protein) SOD (U/mg protein), GSH-Px (U/mg protein), MDA (nmol/mg protein), Insulin (μU/mg protein) and serum NO (μmol/l) of all groups, A (control), B (STZ-treated) and C (STZ- +AME-treated) rats.
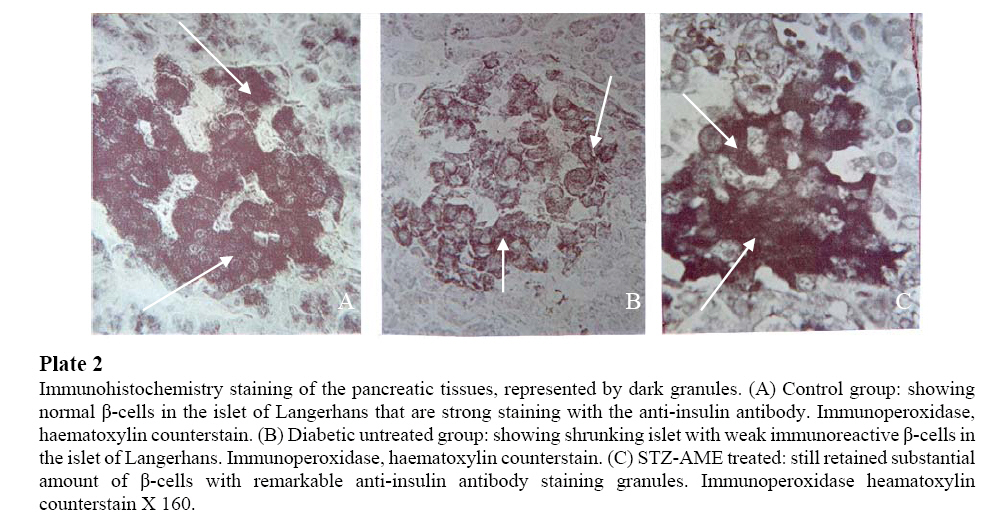
Values are expressed as means (±SEM) of 10 rats for all groups. a Insignificant difference (p>0.05) between all groups. b Significant difference (p<0.05) in the same row between treatment and control groups. Furthermore, high levels of blood glucose concentrations of the STZ-treated rats were persistently observed throughout the study period (21.6±0.3 mmol/l). The blood glucose concentrations in the AME-treated group, (5.6 mmol/l) significantly reduced in value (p < 0.05). Biochemical findings: Table 3 shows the effects of A. muricata on biochemical variables suggestive of oxidative stress in STZ-treated animals. There was clear evidence that STZ-induced pancreatic injury was associated with free radical injury and oxidative stress. Oxidative stress was characterized by increased lipid peroxidation and/or altered non-enzymatic and enzymatic antioxidant systems (Zima et al; 2001). Effects of STZ and STZ + AME treatments on pancreatic tissue’s CAT, GSH, SOD, GSH-Px, MDA, insulin and serum NO are presented in Table 3. The pancreatic antioxidant activity of CAT, GSH, SOD, GSH-Px and pancreatic insulin significantly decreased (p<0.05), while pancreatic MDA and serum NO significantly increased in the STZ-treated, diabetic group of rats. The control group rats maintained optimal value activity of the antioxidants studied. Annona muricata treatment significantly (p<0.05) decreased the elevated NO and MDA, but also significantly increased (p<0.05) the reduced antioxidant enzyme activities. Also, AME proved significantly better in restoring the altered activity of antioxidant enzymes like CAT, GSH, SOD, GSG-Px and MDA, NO and insulin towards their normal values in the pancreas. Histopathological findings Histologically, in diabetic rats with no treatment, the most consistent findings in the sections of pancreatic tissues were breakdown of micro-anatomical features such as extensive β-cell degranulation, decreased cellular density, and an indistinct boarder between the endocrine and exocrine regions. Also there was a diffused degenerative and necrotic changes, and shrunken in the islet of Langerhans (Plate 1b). The nucleus of necrotic cells showed either pyknosis of marginal hyperchromasia, and the cytoplasm were filled by hydropic changes (Plate 1b). The α-cells, exocrine pancreatic acinar epithelium, ductal and connective tissues appeared normal (Plate 1a and b). Identifiable pancreatic islets of diabetic rats as from day 10 were of low cellularity, although there was no evidence of inflammatory cell infiltration (Plate 1b). In diabetic rats treated with A muricata, there was a remarkable improvement in the islet of Langerhans with distinct cellularity changes, majority of the cells showed viable islet of Langerhans, with increase in granulation (Plate 1a). Immunohistochemical findings Immunohistochemistry staining of the pancreatic tissues before STZ injection showed the presence of a strong islet insulin immunoreactivity at a level of 0.59 islet/mm2 of total pancreatic tissue. This was limited to cytoplasmic staining of individual β-cells (Fig. 4A). The majority of islets from AME-treated pancreases stained positive for insulin suggesting that the architecture of AME-treated rats was normal (Fig. 4C). In contrast, islet cells from diabetic rats were architecturally distorted, containing significantly fewer insulin-positive cells (Fig. 4B). Quantitative image analysis was used to assess the proportion of insulin positive cells per islet in control, STZ-treated and STZ + AME-treated pancreatic sections. The percentage of islets stained for insulin were 76.35 ± 19%, 42.26 ± 3.2% and 68.56 ± 24% (p < 0.05) respectively (Fig. 4A, 4B and 4C). Together, these findings further support that AME protected islets from destruction. Of interest is the stability of pancreatic insulin content of the AME-treated rats (15.6±2.1 μU/mg), in contrast with the STZ-treated rats, pancreatic insulin contents were ~65-fold higher (Table 2). This stability in pancreatic insulin values was also observed to reflect in the immunohistochemistry staining (Fig. 4C). Immunohistochemistry staining intensity was an evidence of protective effects/or regenerative processes leading to increase in β-cell mass, regaining its normal immunostaining for insulin and functional status up to the day of normoglycemia. DISCUSSION The aim of the present study was to demonstrate the efficacy of Annona muricata in the reduction of blood glucose concentration as well as to determine the recovery in altered biochemical variables, indicative of oxidative stress and organ damage in STZ-treated rats. Ethnobotanically, all parts of A. muricata tree have been used medicinally in the tropics, including the bark, leaves, roots, fruit, and fruit seeds (Mors, 2000; Haddock, 1994). Different properties and uses are attributed to the different parts of the A. muricata by the Traditional herbal medicine practitioners (Padma et al; 2001). Generally, the fruit and fruit juice are taken for worms and parasites, to cool fevers, to increase mother’s milk after childbirth, and as an astringent for diarrhea and dysentery. The crushed seeds are used against internal and external parasites, head lice and worms. The bark, leaves, and roots are considered sedatives, antispasmodic, hypoglycemic, hypotensive, diuretics, neuralgia (Hasrat et al; 1997). Phytochemically, many active compounds and chemicals have been found in A. muricata, as scientists have been studying its properties since the 1940s. The tree is rich in ellagic acid, tannins, lactones, and isoquinoline alkaloids. The leaf, stem, bark and seeds also contain varying amounts of a novel group of chemicals believed to be biologically active, called Annonaceous acetogenins (Tormo et al; 2003; Kojima, 2004), and this include: annocatalin, annohexocin, annomonicin, annomontacin, annomuricin, annomutacin, annonacin to mention a few. Purdue researchers reported that 14 different acetogenins tested thus far demonstrate potent ATP-blocking properties and were more potent against multi-drug resistant (MDR) cancer cells (Oberlies et al; 1995). A tumor cell needs energy to grow and reproduce and by inhibiting energy to the cell, it can no longer run its pump and expel attacking agents (Yuan et al; 2003). The increasing prevalence of diabetes is reaching epidemic proportion worldwide. Diabetes is a major threat to global public health that is rapidly escalating (WHO, 2000). It is estimated that more than 170 million people are suffering from diabetes globally and this number is expected to double by 2030, and the greatest increase in prevalence is, however, expected to occur in Asia and Africa, follow the trend of urbanization and lifestyle changes (WHO, 1999). Diabetes and its multiple complications are extremely burdensome on the health and economics of countries worldwide. In developing countries adherence to therapies is as low as 20%, resulting in poor health outcomes at a very high cost to society, governments and families (Caro et al; 1987). If not successfully managed, diabetes along with other chronic diseases will become the most expensive problem faced by the health care systems. Because of its associated morbidity and mortality, it is exerting a major pressure on the healthcare system, and with a better understanding of the pathophysiology, a new care perspective and reform in the training of the health workforce is essential. The new care perspective advocates for the use of the Innovative Care for Chronic Conditions framework which integrates prevention, control and treatment of the disease across multiple levels of health care system particularly in low resources settings. In Africa, many plants have been reported to possess antidiabetic or hypoglycemic activities (Ojewole, 2001; Iwu, 1993; Marles and Farnsworth, 1995). Indeed, hundreds of plants are now being used traditionally for the management of both insulin-dependent (IDDM) and noninsulin-dependent (NIDDM) diabetes mellitus. To date, however, only a few of these plants have been subjected to scientific scrutiny either in laboratory animals or in human subjects (Ross, 1999). Although diabetes has been recognized since antiquity, and treatments of various efficacy have been known since the middle ages, the elucidation of the pathogenesis of diabetes occurred mainly in the 20th century (Nathan et al; 2005). The discovery of the action and importance of insulin in the metabolism of glucose, not surprisingly, led to the belief that lack of insulin was the critical factor in the development of the disease (Patlak, 2002). However, subsequent discovery that insulin administration was not necessary for the reduction of glycosuria and indeed for the control of the disease raised ‘queries’ about the role of insulin. The later demonstration that diabetes had developed in persons whose serum insulin levels were not only adequate, but excessive, further confounded the ideas and thinking of both clinicians and researchers in the field of diabetes. Birth of the concept of insulin resistance then took place and provided an answer to some of the questions. Nevertheless insulin production by the beta cells continued to attract attention and β-cell failure in type 2 diabetes is now being demonstrated (Knowler et al; 2002). What must be of further interest now, however, is concept of incorporation of scientifically proved herbal medicine. The three main classes of biological macromolecules that are susceptible to free radical attack under diabetic condition are lipids, nucleic acids and proteins; the attacks affects the entire tissues in the body and suffer oxidative damage (Halliwell and Gutteridge, 1989). Primary effect of lipid peroxidation is decreased membrane fluidity, which alters membrane properties and can significantly disrupt membrane-bound proteins (Tappel, 1975). The oxidation of proteins includes oxidation of sulfhydryl groups, reduction of disulfides, oxidative adduction of amino acid residues close to metal-binding sites via metal-catalyzed oxidation, reactions with aldehydes, protein-protein cross-linking, and peptide fragmentation (Standman and Oliver, 1991; Starke-Reed and Oliver, 1989). Oxidative damage to nucleic acids includes adducts of base and sugar groups, single- and double-strand breaks in the backbone, and cross-links to other molecules, this causes damage to all four bases and thymine-tyrosine cross-link (Dizdaroglu, 1992; Halliwell and Dizdaroglu, 1992). In the present study we used aqueous extracts, obtained from A. muricata leaves and studied its effect on pancreatic β-cells. Our observations demonstrated that STZ-induced oxidative stress and β-cell damage as revealed by histological and immunohistochemical staining results. Depletion of endogenous antioxidants and generation of free radicals were also noted which are in consonance with the findings of Buttke and Sandstorm (1994). A. muricata caused significant decrease in elevated blood glucose and an increase in serum insulin concentrations as well as pancreatic insulin contents in STZ-treated diabetic rats. Histologically, A. muricata partly protected pancreatic β-cell integrity. It saved the tissue from the usual shrunken islets, denegation, degranulation, hydropic and necrotic changes peculiar to STZ-induced pancreatic injuries. Immunohistochemistry staining was evident of significantly better restoration of altered micro-anatomy of islet of Langerhans of STZ-treated diabetic rats. We also analyzed the intracellular level and activities of GSH, SOD, CAT, GSH-Px, MDA, NO and insulin contents and we found that STZ treatment caused significant increases in blood glucose concentration, MDA and NO generation, and decreased GSH, SOD, CAT, GSH-Px, activities as well as insulin levels when compared with control rats. Decreases in GSH, SOD, CAT and GSH-Px, and simultaneous increases in MDA and NO activities, reflect susceptibility of pancreas to STZ’s significant oxidative stress. Thus, this study has demonstrated that STZ alters the activity of oxidant defenses in the pancreas. This observation is in agreement with the studies reported by Henriksen (2002), Gul et al; (2002) and Derouich and Boutayeb (2002). The data obtained in the present study do not allow definite conclusion to be drawn on the mechanisms of action of AME in the experimental animal paradigms used. However, a number of investigators have shown that tannins and other polyphenolic compounds (e.g., coumarins), flavonoids, triterpenoids, saponins, and a host of other plant secondary metabolites possess hypoglycemic, hypotensive, anti-inflammatory, and other pharmacological properties in various experimental animal models (Ojewole, 2004; 2005; Akah and Okafor, 1992). A. muricata is known to contain ellagic acid, tannis, flavonoids, polyphenolic compounds, β-sistosterol, and catechins (Ames et al; 1993; Decker, 1997; Barch et al; 1994; Alldrick et al; 1986); therefore it is not unreasonable to speculate that some of these chemical compounds especially the coumarins and flavonoids are probably responsible for recovery in the altered biochemical variables and pancreatic tissues. Based on our findings, we conclude that STZ administration is associated with oxidative stress in pancreatic tissues. Nevertheless, A. muricata exhibited antioxidant activity, and is able to diminish and/or prevent, pancreatic oxidative damage produced by STZ. An increased consumption of antioxidants in the diet of individuals is strongly recommended, so that when an individual is subjected to greater oxidative stress, he/she would have better antioxidant defense characteristics, thus counteracting the effects of any pro-oxidant. However, further studies are needed before antioxidants can be used safely as food additives and supplements. Acknowledgements The authors are grateful to Professor J.A.O. Ojewole for his expertise and useful advice in some of the procedures. We also thank Messrs Adeogun Oludele, Doherty O. Wiston for their technical assistance in plants processing. Ms Kogie for her excellent technical assistance in tissue processing. REFERENCES
Copyright 2006 - Ibadan Biomedical Communications Group The following images related to this document are available:Photo images[md06030p2.jpg] [md06030p1.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}