 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
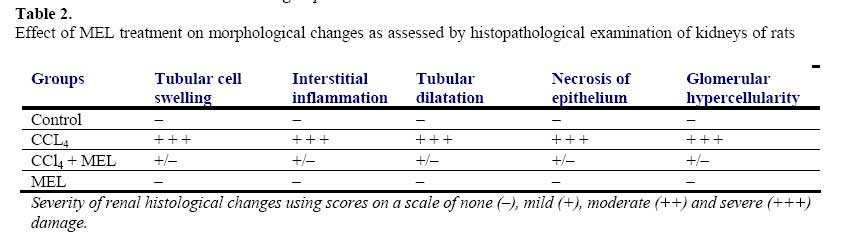
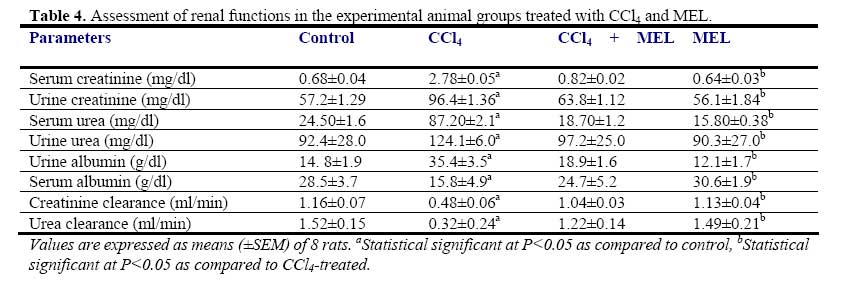
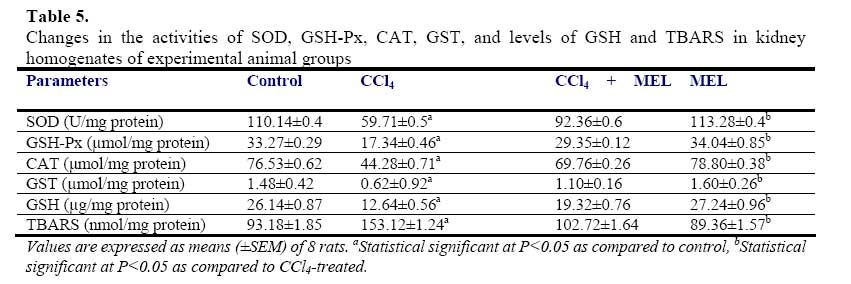
African Journal of Biomedical Research, Vol. 10, No. 2, 2007, pp. 153-164 Effect of Melatonin on Carbon Tetrachloride-Induced Kidney Injury in Wistar Rats Stephen O. Adewole1*, Abdulkadir A. Salako2, Oladepo W. Doherty3 and Thajasvarie Naicker4* Department of 1*Anatomy and Cell Biology, 2Surgery, 3Chemistry Faculty of Basic Medical Sciences, College of Health Sciences; Obafemi Awolowo University, Ile-Ife, Nigeria;4*Optics and Imaging Centre, Nelson R Mandela Medical School, University of KwaZulu-Natal, Private Bag X54001, Durban 4000, South Africa. Received: October 2006 Code Number: md07021 ABSTRACT Exposure to carbon tetrachloride (CCl4) induces acute and chronic renal injuries as well as oxidative stress in rats. The aim of this study was to evaluate the effect of exogenous melatonin (MEL) treatment on CCl4-induced oxidative stress and nephrotoxicity in rats using histopathological and biochemical parameters. Serum creatinine, blood urea nitrogen (BUN), nitrite and albumin concentrations were measured for the evaluation of renal function. Antioxidant status in the kidney tissue was estimated by determining the activities of superoxide dismustase (SOD), glutathione peroxidase (GSH-Px), catalase (CAT) and glutathione-S-transferase (GST) as well as thiobarbituric acid reactive substances (TBARS) and reduced glutathione (GSH) levels. For histopathological evaluation, kidney of all rats were excised and processed for light microscopy. CCl4 caused elevated level of TBARS and marked depletion of renal endogenous antioxidant enzymes. Furthermore, severe deterioration of renal function was observed in CCl4-treated rats as assessed by increased serum creatinine, BUN levels and decreased creatinine and urea clearance as compared with the control rats. MEL treatment positively ameliorated the alterations in these biochemical variables in the CCl4 + MEL-treated rats. MEL markedly reduced elevated TBARS and nitrite, significantly attenuated renal dysfunction, increased the levels of antioxidant enzymes. Kidney sections of CCl4-treated group showed, changes in microanatomy. In contrast, these deleterious histopathological alterations resulting from CCl4 nephrotoxin were absent after MEL treatment in CCl4 + MEL group of rats. In conclusion, our results demonstrated that MEL through its antioxidant activity effectively ameliorated CCl4induced nephrotoxicity. Keywords: Carbon tetrachloride; Melatonin; Nephrotoxicity; Renal dysfunction; Antioxidants INTRODUCTION The pathogenesis of Carbon tetrachloride (CCl4)induced renal dysfunction is not completely known. It may be due to the functional state of liver, or renal injury may develop independently to hepatic events (Rincon et al., 1999). CCl4 induces oxidative stress in many settings (Abraham et al., 1999); therefore, it might be expected to contribute to nephrotoxicity. CCl4 does not occur naturally, it is a clear liquid with sweet smell that can be detected at low levels (Doherty, 2000). Exposure to various compounds including a number of environmental pollutants and drugs can cause cellular damages through metabolic activation of those compounds to a highly reactive oxygen species (ROS). Free radical induced lipid peroxidation is believed to be one of the major causes of cell membrane damage leading to a number of pathological situations (Slater, 1984). CCl4 was formerly used for metal degreasing and as drycleaning, fabric-spotting, and fire extinguisher fluids, grain fumigant and reaction medium. Because of its harmful effects, these uses are now banned and it is only used in some industrial applications (DeShon, 1979). The primary routes of potential human exposure to CCl4 are inhalation, ingestion, and dermal contact. High exposure to CCl4 can cause liver, kidney and central nervous system damage, and liver is especially sensitive to CCl4 because of its role as the body’s principal site of metabolism (Sakata et al., 1987). Reports from our laboratory and other investigators have established that CCl4 is a potent environmental hepatotoxin (Szymonik-Lesiuk et al., 2003). A number of reports clearly demonstrated that in addition to hepatic toxicity, CCl4 also causes disorders in kidneys, lungs, testis as well as in blood by generating free radicals (Ahmad et al., 1987; Ozturk et al., 2003). Findings by Perez et al., (1987), Ogeturk et al., (2005) and Churchill et al., (1983) suggested that exposure to this solvent causes acute and chronic renal injuries. In addition, report on various documented case studies established that CCl4 produces renal diseases in human (Ruprah et al., 1985; Gosselin et al., 1984). A number of endogenous and exogenous nephropathy risk factors generate oxygen free radicals in vivo. Therefore the role of oxygen-derived free radicals and lipid peroxidation has attracted considerable attention (Gebhardt, 2002; Das et al., 2005). Volatile organic compounds such as CCl4 are a class of solvents to which many people are exposed occupationally and environmentally. Early studies of dichloromethane, CCl4 and 1,1-dichloroethylene revealed susceptibility of rats to liver and kidney damage by these chemicals (Bruckner et al., 1984). It has been found that metabolism of CCl4 involves in the production of free radicals through its activation by drug metabolizing enzymes located in the endoplasmic reticulum (Slater and Sawyer, 1971). Therefore, it has become a task to prevent nephropathy and hepatic damage by eliminating free radicals and prevent lipid peroxidation through the use of terminal antioxidant like melatonin (Esrefoglu et al., 2005). Cumulative data suggest a role for reactive oxygen metabolites as one of the postulated mechanisms in the pathogenesis of CCl4 nephrotoxicity (Recknagel et al., 1989). CCl4 results in enhanced generation of trichloromethylperoxyl radical (Cl3COO-), hydrogen peroxide in cultured hepatocytes as well as mesangial cells (Knight et al., 1989). In vitro and in vivo studies indicate that CCl4 enhances lipid peroxidation, reduces renal microsomal NADPH cytochrome P450, and renal reduced/oxidiced glutathione ratio (GSH/GSSG) in kidney cortex as well as renal microsomes and mitochondria (Rungby and Ernst, 1992). Antioxidants such as melatonin, ascorbate, αtocopherol, silibinin, lazaroid, propinonyl carnitine and superoxide dismutase/catalase, have been shown to ameliorate CCl4-induced renal toxicity (Miller and Rice, 1997). Nitric oxide (NO) is a free radical formed from the terminal guanidine group of L-arginine by enzymatic reaction involving NO synthase (NOS). NO is one of the principal mediators involved in the pathogenesis on inflammatory responses partly through direct effects on nuclear factor ĸB (NFĸB). Endogenous inducible NO synthase (iNOS)-derived NO has a protective role in proteinuric tubulointerstitial injury, but excess production has adverse effects in the same situation. Among its multiple divergent functions, NO has several antiinflammatory properties. Apart from maintaining perfusion on the renal microcirculation, NO inhibits leukocytes and platelet endothelial adhesion (Radomski et al., 1987), suppresses T-cell and monocyte proliferation and preserves integrity of the vascular endothelial permeability barrier (Suttorp et al., 1996). NO also is a physiologically important scavenger of superoxide radicals, been more potent than superoxide dismutase (Rubanyi et al., 1991). Mammalian cells are equipped with both enzymic and non-enzymic antioxidants defenses with different efficacies that protect animals against oxidative abuse caused by wide range of nephrotoxicants including CCl4 (Karbownik et al., 2001). Melatonin, N-acetyl-5-methoxytrptamine is a hormone found in all living creatures from algae (Caniato et al., 2003) to humans, at levels that vary in a diurnal cycle. In higher animals MEL is produced by pinealocytes in the pineal gland. Melatonin participates in many important physiological functions, including anti-inflammatory (Cuzzocrea and Reiter, 2002), also, it is a powerful antioxidant that can easily cross cell membranes and the blood-brain barrier (Hardeland, 2005). Unlike other antioxidants, MEL does not undergo redox cycling, once oxidized; it cannot be reduced to its former state because it forms several stable endproducts upon reacting with free radicals (Tan et al., 2000). Redox cycling may allow other antioxidants such as vitamin C to act as pro-oxidants, counterintuitively promoting free radical formation. Furthermore, MEL has been demonstrated to prevent damage to DNA by some carcinogens, stopping the mechanism by which they cause cancer (Karbownik et al., 2001; Carrillo-Vico et al., 2005). Melatonin is an immunomodulatory agent that enhances T-cell production and when taken in conjunction with calcium, it is a potent immunostimulator of the Tcell response (Vijayalaxmi et al., 2002). It holds the unique position of being the only known chronobiotic regulator of neoplastic cell growth. Both in vitro and in vivo, MEL has been found to inhibit neoplastic growth and to delay tumor progression (Tan et al., 1993). In addition, MEL protects liver and kidney injury induced by endotoxin shock and ischemia/reperfusion in rats through its antioxidant action (Sener et al., 2003). Beneficial effects of exogenous MEL on vital organs like brain, liver, kidney and lungs as well as cardiovascular system have been largely reviewed (Zenebe et al., 2001). Therefore, the present study was designed to (i) evaluate beneficial effect of exogenous melatonin in preventing acute renal dysfunction/nephrotoxicity induced by CCl4 in rats, and (ii) analyze degree of kidney injury via antioxidative mechanisms MATERIALS AND METHODS Animal care and monitoring: This study was carried out in healthy, male and female Wistar rats, weighing 240-260 g. The animals were housed under standard laboratory conditions of light, temperature (21±2oC) and relative humidity (55±5%). The animals were given standard rat pellets and tap water ad libitum. The rats were randomly divided into four experimental groups: A (control), B (CCl4), C (CCl4 + MEL) and D (MEL), each group consisted of 10 rats. The experimental protocol and procedures used in this study were approved by the Ethics Committee of the University of KwaZulu-Natal, Durban 4000, South Africa; and conform with the “Guide to the Care and Use of Animals in Research and Teaching” [Published by the Ethics Committee of the University of Durban-Westville, Durban 4000, South Africa]. Experimental design: Animals were divided into four groups of ten rats each. Group A rats were treated with olive oil in quantities equivalent to the volume of CCl4 and MEL administered subcutaneously. The rats in groups B and C were treated with CCl4 (1 ml/kg body weight, s.c.) in a 1:1 solution with olive oil. The rats in group C were additionally treated with MEL (5 mg/kg/, s.c.) for 3 days prior to administration of CCl4; and MEL treatment was continued for further 8 weeks at alternate days. Melatonin was dissolved in (9 g/L) saline with absolute ethanol (oC until used. Group D rats received MEL (5 mg/kg/, s.c.) treatment for a period of 8 weeks, at alternate days. All the animals were fasted for 16 hours, but still allowed free access to water, before the commencement of our experiments. At the end of experimental period of 8 weeks, the animals were sacrificed by cervical dislocation; serum/plasma samples and liver tissue were collected and stored at -80oC for analysis. Kidney histopathology: Kidney tissues were excised from sacrificed animals, individually weighed, and thin kidney slices were cut, fixed in 4% paraformaldehyde and were sequentially embedded in paraffin wax blocks. Tissue sections of 5 μm thick were cut, and stained with hematoxylineosin (H-E) and masson’s trichrome (Suzuki and Suzuki, 1998) for conventional morphological evaluation, then examined under light microscope (BX50; Olympus, Tokyo). The images were obtained by a digital camera system (Pixcera Co., Osaka, Japan) attached to the microscope. A minimum of 10 fields for each kidney slide were examined and scored semiquantitatively for severity of changes by a pathologist unaware of the type of treatment. The scoring was done as none (-), mild (+), moderate (++) and severe (+++). Determination of Creatinine (Cr) and BUN levels and clearances (Ccl ): Before sacrifice, rats were kept individually in metabolic cages (TP-85M; Toyoriko, Tokyo). After 4-5 days adaptations to the cage, urine samples were collected in ice-cooled glass flasks every 8 h for 24 h and stored at 4oC. Urine samples from each rat were combined and measured. After centrifugation, aliquots of the supernatant were frozen at –40oC. Rats were sacrificed under light anesthesia with halothane by decapitation. The abdomen was opened by transverse incision and kidneys were excised for histological studies and enzymatic analysis. Blood samples were collected by cardiac puncture into heparinized container. BUN values as indicators of renal damage were determined by the NADHcoupled enzymatic method, using urase spectrophotometrically in an enzyme-based assay (Hallett and Cook, 1971). Urine and serum samples were assayed for creatinine, using a spectrophotometric assay method of Junge et al., (1998). For the assay, urine was diluted 1:50 in distilled water. Three microliters plasma or diluted urine and 300 µl picric acid reagent (consisting of saturated aqueous picric acid diluted 5-fold in 0.25 M NaOH) were dispensed in duplicate, using a MicroLab dilutor (Hamilton, Reno, NV) into wells of a microtiter plate. After incubation for 15 min at room temperature, absorbancewas read at 515 nm in a microplate reader. Creatinine clearance was calculated according to Cockcroft-Gault formular (Cockcroft and Gault, 1976). Ccr = (Cu/Cs) × V, where Cu is the concentration of creatinine in urine, Cs is the concentration of creatinine in serum and V is the urine flow rate in milliliters per minute. Serum and Urine albumin concentration: Serum albumin concentration was measured in a multistat III Plus machine with reagents from Fisher Scientific (St. Louis, MO). Urine samples were collected into test tubes and stored at –40oC for analysis. The concentration of urinary albumin was determined using standard diagnostic kits, an enzyme-linked immunosorbent assay (ELISA) (Nephrat Exocell Inc., Philadelphia, PA, USA). Assessment of urine/serum and tissue nitrite concentration: Urine samples were deproteinized by equal volumes of 0.3M NaOH and 5% zinc sulphate and centrifuged at 6400 x g for 20 min. The supernatants were added in duplicates to 96-well plates and reacted with Greiss reagent (1:1 solution of 1% sulphanilamide in 5% phosphoric acid and 0.1% napthaylamine diamine dihydrochloric acid in water). Also serum and tissue nitrite was estimated using Greiss reagent and served as an indicator of NO production. 500 µl of Greiss reagent was added to suitably diluted 100 µl of serum and supernatant of the tissue as described above and corrected by protein amount. Nitrite levels were assessed by measurement of absorbance at 540 nm using a micro plate reader (Model 550; Bio-Rad laboratories, Hercules, CA) (Green et al., 1982). Nitrite concentration was calculated using a standard curve for sodium nitrite. Nitrite levels were expressed as µmol/ml in serum and urine and as µmol/mg protein in homogenate. Preparation of tissue homogenate: The harvested kidney tissues were rinsed in cold physiological saline, cleaned of gross adventitial tissue, blotted dry and stored in a Biofreezer at –80oC until analyzed. Liver tissue was homogenized with Potter Elvehjem homogenizer. 10% homogenates were prepared in 6.7 mM phosphate buffer, pH 7.4 and centrifuged at 10,000 rpm for 10 min at 4oC, and the resultant supernatant was used for measurement of antioxidant enzymes. For the determination of lipid peroxidation, liver tissue was homogenized in 1.15% KCl solution to obtain a 10% (w/v) homogenate. Protein content of liver homogenates was determined by using bicinchoninic acid (BCA) protein assay reagent (Pierce Chemical Company, Rockford, IL). All enzyme activities were expressed as units/mg protein. Biochemical assays: Superoxide Dismutase Activity (SOD): Kidney SOD activity was assayed by the method of Kakkar et al., (1984). Reaction mixture contained 1.2 ml of sodium pyrophosphate buffer (0.052 mM, pH 7.0), 0.1 ml of phenazine methosulphate (PMS) (186 μM), 0.3 ml of nitro blue tetrazolium (NBT) (300 μM). 0.2 ml of the supernatant obtained after centrifugation (1500 x g, 10 min followed by 10,000 x g, 15 min) of 10% kidney homogenate was added to reaction mixture. Enzyme reaction was initiated by adding 0.2 ml of NADH (780 μM) and stopped precisely after 1 min by adding 1 ml of glacial acetic acid. Amount of chromogen formed was measured by recording color intensity at 560 nm. Results are expressed as units/mg protein. Glutathione Peroxidase Activity (GSH-Px): Glutathione peroxidase (GSH-Px) activity was measured by NADPH oxidation, using a coupled reaction system consisting of glutathione, glutathione reductase, and cumene hydroperoxide (Tappel, 1978). 100 µL of enzyme sample was incubated for five minutes with 1.55 ml stock solution (prepared in 50 mM Tris buffer, pH 7.6 with 0.1 mM EDTA) containing 0.25 mM GSH, 0.12 mM NADPH and 1 unit glutathione reductase. The reaction was initiated by adding 50 µL of cumene hydroperoxide (1 mg/ml), and the rate of disappearance of NADPH with time was determined by monitoring absorbance at 340 nm. One unit of enzyme activity is defined as the amount of enzyme that transforms 1 µmol of NADPH to NADP per minute. Results are expressed as units/mg protein. Catalase Activity (CAT): The activity of CAT was measured using its perioxidatic function according to the method of Johansson and Borg, (1988). 50 µL potassium phosphate buffer (250 mM, pH 7.0) was incubated with 50 µl methanol and 10 µL hydrogen peroxide (0.27%). The reaction was initiated by addition of 100 µL of enzyme sample with continuous shaking at room temperature (20oC). After 20 minutes, reaction was terminated by addition of 50 µL of 7.8 M potassium hydroxide. 100 µL of purpald (4-Amino-3-hydrazino-5mercapto-1,2,4-triazole, 34.2 mM in 480 mM HCl) was immediately added, and the mixture was again incubated for 10 minutes at 20oC with continuous shaking. Potassium peroxidate (50 µL 65.2 mM) was added to obtain a colored compound. The absorbance was read at 550 nm in a spectrophotometer. Results are expressed as micromoles of formaldehyde produced/mg protein. Reduced glutathione (GSH): Reduced glutathione (GSH) level in the kidney was assayed following the method of Ellman (1959), modified by Hissin and Hilf (1973). The homogenate (720 µl) was double diluted and 5% TCA was added to it to precipitate the protein content of the homogenate. After centrifugation (10, 000 x g for 5 minutes) at 4oC the supernatant was taken, 5,5´-dithiolbis-2-nitrobenzoic acid (DTNB) solution (Ellmam’s reagent) was added to it and the absorbance was measured at 412 nm on a spectrophotometer. A standard graph was drawn using different concentrations of standard GSH solution (1 mg/ml). With the help of the standard graph, GSH contents in the homogenates of the experimental animals were calculated. Glutathione-S-transferase (GST): GST catalyzes the conjugation reaction with glutathione in the first step of mercapturic acid synthesis. GST activity was measured by the method of Habig and Jakoby (1974). The reaction mixture contained suitable amount of the enzyme (25 µg of protein in homogenates), 1 ml of KH2PO4 buffer, 0.2 ml of EDTA, 0.1 ml of 1-chloro-2,4-dinitrobenzene (CDNB), and GSH. The reaction was carried out at 37oC and monitored spectrophotometrically by the increase in absorbance of the conjugate of GSH and CDNB at 340 nm. A blank was run in absence of the enzyme. One unit of GST activity is 1 µmol product formation per minute. Thiobarbituric Acid Reactive Substances (TBARS): The product of the reaction between malondialdehyde (MDA) and thiobarbituric acid reactive substances (TBARS) were measured by the method of Ohkawa et al., (1997). The reaction mixture consisted of 0.2 ml of 8.1% sodium lauryl sulphate, 1.5 ml of 20% acetic acid solution adjusted to pH 3.5 with sodium hydroxide and 1.5 ml of 0.8% aqueous solution of thiobarbituric acid was added to 0.2 ml of 10% (w/v) of homogenate. The mixture was brought to 4.0 ml with distilled water and heated at 95oC for 60 minutes. After cooling with tap water, 1.0 ml distilled water and 5.0 ml of the mixture of n-butanol and pyridine (15:1 v/v) was added and centrifuged at 3000 rpm for 10 minutes. The organic layer was taken out and absorbance of the clear upper (n-butanol) layer was measured using Shimadzu UV-1601 (Japan) spectrophotometer at 532 nm. TBARS were quantified using an extinction coefficient of 1.56 x 105 cm-1 M-1 and expressed as nmol of TBARS/mg tissue protein. Tissue protein was estimated using Biuret method of protein assay and the renal MDA content expressed as nmol of MDA per mg protein. Statistical Analysis: The data obtained were expressed as means (±SEM). The inter-group variation was measured by one way analysis of variance (ANOVA; 95% confidence interval) followed by Fischer’s LSD test. Statistical significance was considered at p<0.05. RESULTS Body and kidney weights: Table 1 shows the changes in body and kidney weights of all experimental animal groups. Administration of CCl4 significantly increased (p<0.05) kidney weight and at the same time significantly decreased body weight when compared with the control and MEL-treated groups of rat. MEL treatment significantly (p < 0.05) reduced relative kidney weight, and produced a statistically near equal (p = 248.3) body weight with the body of control rats. The CCl4 + MEL group insignificantly showed changes in kidney and body weights (Table 1). Histopathological findings: CCl4 administration alone caused prominent histopathological damage in the kidney compared with the control rats. The histopathological changes were graded and summarized (Table 2). The kidney sections from the control- and MEL-treated groups showed normal glomeruli, and tubulointerstitial cells (Plate 1A and 1D). In contrast, the kidneys of CCl4-treated rats showed marked deleterious histological changes. The kidney sections showed significant glomerular and tubular degenerations varying from, glomerular basement thickening, interstitial inflammation, tubular cell swelling, pycnotic nuclei, medullary vascular congestion and moderate to severe necrosis (Plate 1B). MEL-treated group of rats preserved normal morphology of the kidney and shows normal architecture of the kidney (Plate 1C). Urine/Serum and tissue nitrite levels: Administration of CCl4 significantly increased (p<0.05) urine, serum and tissue nitrite levels in CCl4-treated rats. Melatonin treatment significantly decreased (p<0.05) the elevated nitrite levels in urine, serum and tissue of CCl4 +MEL-treated rats. However MEL per se has no effect on urine, serum and tissue nitrite levels of group D rats (Table 3). Serum creatinine, BUN and albumin levels: CCl4 treatment significantly increased (p<0.05) the serum creatinine, BUN and urine albumin levels as compared with the control group of rats. However, serum albumin levels were significantly lower in CCl4-treated rats (Table 4). Although, treatment of rats with MEL significantly prevented this rise in serum creatinine, BUN and urine albumin and at the same time increased serum albumin in CCl4 + MEL group of rats (Table 4). Creatinine and urea clearance, which was markedly decreased in CCl4treated rats, was significantly increased by MEL treatment (Table 4). MEL treatment per se, however, had no effect on these biochemical variables in group D rats (Table 4). Biochemical findings: Table 5 shows the effects of melatonin on biochemical variables in the tissue homogenates of all experimental animal groups. CCl4 administration markedly decreased antioxidants enzymes as well as glutathione levels and this is suggestive of oxidative stress in CCl4treated rats. There was clear evidence that CCl4induced renal injury was associated with free radical injury and oxidative stress as evidenced by the activities antioxidants activities (Table 5). Oxidative stress was characterized by increased lipid peroxidation and/or altered non-enzymatic and enzymatic antioxidant systems. Effects of CCl4 and CCl4 + MEL treatments on renal tissue’s SOD, GSH-Px, CAT, GST, GSH and TBARS are presented in Table 5. The renal antioxidant activity of SOD, GSH-Px, CAT, GST and GSH significantly decreased (p<0.05), while renal TBARS significantly increased in the CCl4 - treated, group of rats. The control group of rats maintained optimal value activity of the antioxidants studied. Administration of melatonin significantly (p<0.05) decreased the elevated TBARS, and also significantly increased (p<0.05) the reduced antioxidant enzyme activities. Furthermore, MEL proved significantly better in restoring the altered activity of antioxidant enzymes like SOD, GSG-Px, CAT, GST, GSH and TBARS towards their normal values in the kidney homogenates. The animals treated with MEL alone showed no significant change in the levels of GSH and in the activities of GST related enzymes (Table 5). DISCUSSION In vitro and in vivo studies indicate that CCl4 enhances lipid peroxidation, reduces renal microsomal NADPH cytochrome P450, and renal reduced/oxidized glutathione ratio (GSH/GSSG) in kidney cortex as well as renal microsomes and mitochondria (Walker et al., 1996). In general, a number of chemicals including various environmental toxicants and even clinically useful drugs can cause severe cellular damages in different organs of the body through metabolic activation to highly reactive substances such as free radicals (Noguchi et al., 1982). CCl4 is one of such widely used environmental toxicant to experimentally induce animal models of acute nephrotoxicity and hepatic damages. CCl4 is metabolized by cytochrome P450 2E1 to trichloromethyl radical (CCl3 -). CCl3- and its highly reactive derivative, the trichloromethylperoxyl radical (Cl3 COO-), are assumed to initiate free radical-mediated lipid peroxidation leading to accumulation of lipid peroxidation products that causes renal and hepatic injuries (Aleynik et al., 1997). These radicals are capable of initiating a chain of lipid peroxidation reactions by abstracting hydrogen from polyunsaturated fatty acids (PUFA). Peroxidation of lipids, particularly those containing PUFA, can dramatically change the properties of biological membranes, resulting in severe cell damage and play a significant role in pathogenesis of diseases (Aleynik et al., 1997). This phenomenon results in the generation of ROS, (like superoxide anion O2 -, H2 O2 and hydroxyl radical OH-). Evidence suggests that various enzymatic and non-enzymatic systems have been developed by mammalian cells to cope with ROS and other free radicals (Recknagel et al., 1989). However, when a condition of oxidative stress establishes, the defense capacities against ROS becomes insufficient (Halliwell and Gutteridge, 2000). ROS also affects the antioxidant defense mechanisms, by reducing the intracellular concentration of GSH and decreases the activity of SOD, CAT and GSH-Px. It has also been observed to decrease the detoxification system produced by GST (Yamamoto and Yamashita, 1999). In the kidney GST is a cystolic protein, highly specific for the cells of the proximal tubules. It is found in high concentrations and is readily released into the urine in the event of renal tubular injury (Harrison et al., 1989). Oxidative stress can promote the formation of a variety of vasoactive mediators that can affect renal function directly by initiating renal vasoconstriction or decreasing the glomerular capillary ultrafiltration coefficient; and thus reducing glomerular filtration rate (Garcia-Cohen et al., 2000). Thus the attenuation of lipid peroxidation in CCl4 -treated rats by MEL provides a convincing evidence for the involvement of ROS in CCl4 -induced lipid peroxidation. It is a known fact that MEL, the chief secretory product of the pineal gland, was found to be a multifaceted free radical scavenger and antioxidant. It detoxifies a variety of free radicals and reactive oxygen intermediates, including the hydroxyl radical, singlet oxygen peroxynitrite anion and nitric oxide (Tan et al., 2002). In both in vitro and in vivo experiments, MEL has been found to protect cells, tissues, and organs against oxidative damage induced by a variety of free-radical-generating agents and processes, such as the CCl4 , ischemia reperfusion, amyloid-protein, carcinogen safrole and ionizing radiation (Marchiafava and Longoni, 1999). Melatonin also has been reported to stimulate the activities of enzymes and increase gene expression that improves the total antioxidative defense capacity of the organism such as SOD, GSH-Px, and GSH (Kotler et al., 1998). Recent studies indicate that MEL is effective on inhibiting oxidative and kidney and liver damage (Calvo et al., 2001). Melatonin also could dose-dependently reduce kidney lipid peroxide content in CCl4 -treated rats. This indicated that MEL exerted a therapeutic effect on CCl4 -induced acute or chronic renal injury in rats, possibly through its antioxidant action. Melatonin plays a cytoprotective role in the kidney insulted by ischemia and reperfusion by virtue of its ability to prevent kidney malfunction and inhibit the generation of free radicals and accumulation of neutrophils in the damaged renal tissue (Lepay et al., 1985). From the present study, it has been observed that CCl4 induced significant decrease in SOD, CAT, GSH-Px and GST activities, depleted the GSH content and enhanced lipid peroxidation in kidney. It has been reported that SOD, CAT, GSH-Px and GST constitute a mutually supportive team of defense against ROS (Bandhopadhy et al., 1999). The decreased activity of SOD in kidney in CCl4 treated rats may be due to the enhanced lipid peroxidation or inactivation of the antioxidative enzymes. This would cause an increased accumulation of superoxide radicals, which could further stimulate lipid peroxidation. GST bind to liophilic compounds and acts as an enzyme for GSH conjugation reactions (Anadan et al., 1999). Decreased GST activity during CCl4 toxicity might be due to the decreased availability of GSH resulted during the enhanced lipid peroxidation. Administration of MEL prior to CCl4 intoxication protected the antioxidant machineries of the kidney as revealed from enhanced levels of SOD, CAT, GSH-Px and GST activities, increased GSH content and decreased lipid peroxidation. NO production has been reported to increase in renal disease (Cattell et al., 1990), our results confirm these findings. Nitrite excretion in urine was increased in CCl4 -treated rats and significantly diminished by MEL treatment. In addition, decreased nitrite content in kidney tissue was observed to be significant. However, increased NO in urine might be due to an increased NO production in the glomeruli as a result of changes in L-arginine utilization or an additional activation of systemic macrophages. Also it is possible that NO directly mediates hyperfilteration through its potent vasodilatory properties which is in agreement with findings of Reyes et al. (1992). The present study revealed that chronic administration of CCl4 caused marked impairment in renal function alongside with significant oxidative stress in the kidney. Serum creatinine and BUN concentrations were significantly higher in CCl4 treated rats which are consistent with lower creatinine and BUN clearance. MEL significantly improved creatinine and BUN clearance, and decreased the elevated levels of serum creatinine and BUN. In addition, elevated level of urinary albumin and reduced level of serum albumin concentrations in CCl4 -treated rats might have resulted from remarkable leakage due to hypercellularity of both glomeruli and tubules (Fig. 4B). Histopathological alterations common to CCl4 -treated rats were glomerular hypercellularity, moderate to severe necrosis and tubulointerstitial alterations. It is believed that the capacity for tubular absorption may have been altered, thus bringing about functional overload of nephrons with subsequent renal dysfunctions (Fig. 4B). On the other hand, MEL protect kidney tissue against oxidative damagesinduced by CCl4 In conclusion this study demonstrates that melatonin through its marked antioxidant activity coupled with favourable anti-inflammatory effects salvages CCl4 -induced nephrotoxicity. ACKNOWLEDGEMENTS The authors are grateful to Mr. O. Adeogun, Mr. J Ibeh and Miss K. Moodley for their technical assistance in tissue processing. REFERENCES
© Ibadan Biomedical Communications Group The following images related to this document are available:Photo images[md07021t2.jpg] [md07021t5.jpg] [md07021t3.jpg] [md07021t4.jpg] [md07021t1.jpg] [md07021f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}