 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Middle East Fertility Society Journal, Vol. 9, No. 2, 2004, pp. 163-170 Histological evaluation and In situ localization of apoptosis in fresh and cryopreserved ovarian tissue Mohamed A. Bedaiwy, M.D.*† ,Mahmoud R. Hussein, M.D., Ph.D.‡ Departments of Obstetrics and Gynecology, Minimally Invasive Surgery Center, Cleveland Clinic Foundation, Cleveland, Ohio; department of Obstetrics and Gynecology, and Pathology Department, Assiut School of Medicine, Assiut, Egypt. *Department of Obstetrics and Gynecology, Minimally Invasive SurgeryCenter,
Cleveland Clinic Foundation, Cleveland, Ohio44195 Presented in part in: 58th Annual Meeting of the American Society for Reproductive Medicine, Oct 12-17, 2002Seattle Correspondence: Mohamed A. Bedaiwy, MD, Department of Ob. Gyn., Assiut School of Medicine, Assiut, Egypt, Phone: 088-364258, Fax: 088-333327, Email: bedaiwymmm@yahoo.com Received on May 9, 2004; Code Number: mf04028 ABSTRACT Objective:To study the feasibility of using combined morphology and
terminal deoxynucleotidyl transferase-mediated dUTP-digoxigenin nick-end labeling
(TUNEL) for apoptosis detection and the impact of cryopreservation on this
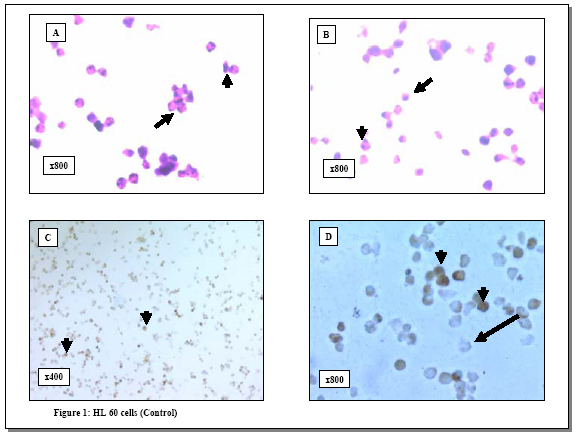
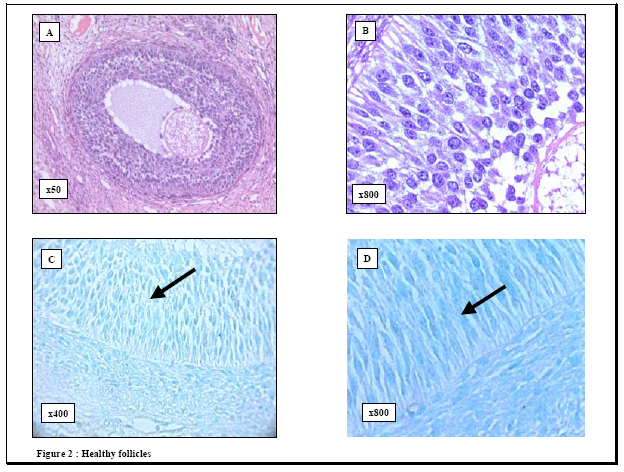
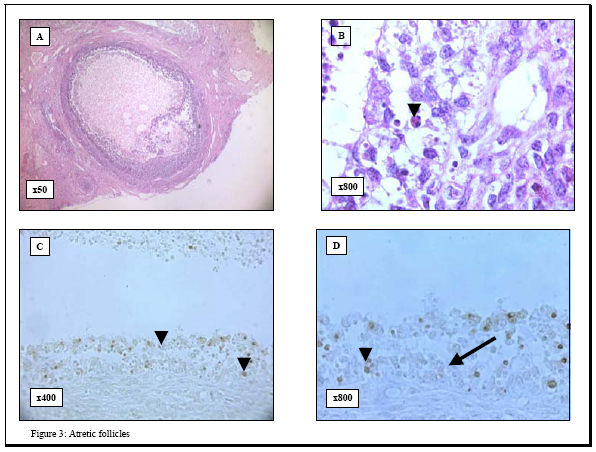
process. Key words: Apoptosis/ovarian cryopreservation/Morphology/TUNEL assay. In 1885, the first observation of apoptosis in the ovary was made following morphological analysis of granulosa cells in the rabbit ovary (1). This form of cell death is involved in the ovarian homeostasis (2) and its regulation depends on a balance between survival factors and apoptotic factors (3, 4). Morphologically, apoptosis is characterized by specific changes, such as cytoplasmic vacuolization, chromatin condensation and formation of apoptotic bodies (5, 6). Biochemically, apoptosis is characterized by DNA fragmentation into oligonucleosome-sized fragments (6). This DNA fragmentation has been examined in the pig ovarian tissues; using DNA fluorescence flowcytometry (7) and autoradiography (8-10). However, these methods have the following limitations 1) they require large amounts of DNA and 2) they do not allow identification of apoptotic changes in specific tissue compartments or at the single cell level. Although morphology provided the original criteria for apoptosis and therefore remains as a reliable indicator, detection of early apoptotic changes may be beyond the scope of histological examination. Alternatively, it is well known that apoptosis may occur without evidence of DNA fragmentation (11) and therefore its detection becomes beyond the scope of terminal deoxynucleotidyl transferase-mediated dUTP-digoxigenin nick-end labeling (TUNEL) assay. The use of morphology in combination with TUNEL assay to examine apoptosis may produce more clear and unequivocal results (12). However, knowledge about this combined strategy is scarce so far. With the growing interest in ovarian tissue cryopreservation to scavenge the fertility of those at risk, the response of the different ovarian cellular elements to the cryopreservation insults is not well studied. To fill this gap in the literature, we examined apoptosis in porcine ovarian tissues using this combined method. We addressed three questions 1) what are the patterns of apoptosis in the pig ovary? Does it affect healthy follicles?; 2) what are the feasibility and advantages of this method?, and 3) what is the effect of cryopreservation on this process? MATERIALS AND METHODS Tissues Bilateral oophorectomy was performed in eight sows at the Cleveland Clinic Foundation Biological Research Unit in accordance with standard operating procedures. Each specimen was divided into two parts; one part was immediately fixed in Bouin solution for histology while the other was cryopreserved according to the protocol by Gosden and his colleagues (13). The cryopreserved tissues were kept in liquid nitrogen for 3 months, and then thawed. Serial 4-6 mm sections were cut from both freshly fixed and cryopreserved tissues, mounted on glass slides and placed in a 60o C oven for 30 minutes. The Bouin-fixed sections were then deparaffinized in xylene, rehydrated through graded alcohol and stained with hematoxylene and eosin for histological examination. Histological evaluation of follicular health and atresia The follicles were categorized into two groups, healthy and atretic, following the established morphological criteria (14). The healthy follicles were those with intact membrana granulosa and few pyknotic nuclei (<5% pyknotic nuclei) in this layer. Alternatively, atretic follicles showed attenuated membrana granulosa, disruption, loosely attached granulosa cells and increased number of pyknotic nuclei (>5% pyknotic nuclei) (15). Histological evaluation of apoptosis Hematoxylene and eosin sections were examined, and histological evaluation of apoptosis followed the established criteria reported in the original paper by Kerr and his colleagues (16). These criteria include condensed nuclear fragments, nuclei with marginated chromatin, multiple nuclear fragments, a single condensed nucleus, membrane-bound structures containing variable amounts of chromatin and/or cytoplasm and eosinophilic cytoplasm. TUNEL assay for detection of apoptotic cells To evaluate the apoptotic response in the pig ovarian tissue, we applied terminal deoxynucleotidyl transferase (TdT)-mediated dUTP-digoxigenin nick-end labeling (TUNEL assay) technique, to both the cryopreserved and Bouin-fixed sections, using the commercially available QIA33TDT-FragELTM kits (Oncogen Research Products, Boston, MA02118, USA). The Bouin-fixed sections (4-5 mm) mounted on glass slides were deparaffinized, rehydrated through graded alcohols to water, treated with 20 µg/ml proteinase K (37o C, 20 minutes) and then washed in 1X Tris buffer. TUNEL assay was then performed according to the instructions by the manufacturer. Briefly, endogenous peroxidase activity was blocked with 3% H202 in methanol. Sections were then rinsed in 1X Tris buffer and covered with 5X TdT equilibration buffer at 37o C for 30 minutes. The buffer was blotted from the slides, and TdT labeling reaction mixture (57.0 µl TdT labeling solution and 3.0 µl TdT enzyme) was added at 37o C for 1.5 hours. The reaction was terminated by rinsing in 1X Tris buffer and covering the sections with 100 µl STOP solution for 5 minutes. Then the sections were incubated with blocking buffer and 50X conjugate solution for 10 and 30 minutes, respectively. The slides were rinsed in 1X Tris buffer, and the entire specimen was covered with DAB solution for 15 minutes. The nuclei were counterstained with 5% methyl green, and the sections were treated with 100% ethanol, cleared in xylene and cover-slipped. Evaluation of apoptosis (TUNEL assay) In accordance with other groups (17, 18), the results of the TUNEL assay were evaluated according to the signal intensity as follows -, negative and +, positive. Positive and negative controls Positive controls were obtained from the manufacturer (Oncogen Research Products, Boston, MA02118, USA) and consisted of formaldehyde-fixed, paraffin-embedded sections from HL60 promyelocytic leukemia cells and HL60 cells incubated with 0.5 µg/ml actinomycin D for 19 hours to induce apoptosis. Some ovarian tissue specimens were used as negative controls by substituting a microliter of distilled water for the deoxynucleotidyl transferase from the protocol, as suggested by others (19-22). All the sections were then examined with a BH2 Olympus microscope. Photography All photographs were taken using a camera attachment with the Olympus BH2 microscope. Statistical analysis Statistical analysis was done using Analysis of Variance (ANOVA) (Statistix for Windows, 1985, 96 Analytical Software Program). Differences were considered statistically significant at p<0.05. RESULTS Histological evaluation When the results between the cryopreserved and freshly- fixed groups were compared: 1) the number of primordial follicles was significantly reduced (4.9±5.3 vs. 7.2±5.4, p=0.03) and 2) the differences in the percentage of the primary and secondary follicles between the two groups were not significant. Generally, granulosa cells with the histological features of apoptosis were observed in atretic follicles either within the central region of the membrana granulosa layer or loosely attached to the membrana granulosa near its antral surface or in the antral follicular fluid (Figures 1, 2, 3). TUNEL assay None of the negative controls stained with TUNEL technique showed any immunoreactivity, while signal-positive TUNEL staining was observed in the positive controls (Figure 1). The evaluation of apoptosis in ovarian tissues revealed several observations. First, the healthy (primordial, primary and secondary) follicles were TUNEL-negative (Figure 2). Second, TUNEL-positive signals were detected in the atretic follicles as indicated by the punctuated brown staining of the fragmented nuclei of granulosa cells (Figure 3). As in histological evaluation, the positively stained nuclei were observed either in the central layers of the membrana granulosa, at the antral surface or floating in the follicular antrium. Apoptosis was absent both in the interstitial cells and in the theca cells of the atretic follicles. Third, although many of the pyknotic nuclei and the DNA globules sloughed in the cavities of the atretic follicles were stained intensely with hematoxylene, they were TUNEL-negative. Forth, although the mean values of apoptotic cells were slightly higher in the cryopreserved tissues as compared to the freshly fixed ones (1.8 vs. 1.8; 2.0 vs. 2.0; 2.1 vs. 2.1; 2.3 vs. 2.5 and 2.4 vs. 2.8 at 1, 5, 10, 20 and 30 minutes respectively), none of these differences were statistically significant. Finally, the localization of apoptosis was not significantly affected by cryopreservation of the ovarian tissue. DISCUSSION Although previous studies have reported apoptotic changes in the pocrine ovarian tissues using several molecular techniques (7-10), these methods require large amounts of DNA and do not allow identification of apoptotic changes in specific tissue compartments or at the single cell level. Alternatively the combined approach proposed by our study has the following advantages 1) it can be performed on conventional histological sections without disruption of their architecture; 2) it allows direct microscopic visualization and detection of apoptosis in specific tissue compartments and at the single cell level; and 3) it is a rapid non-radioactive method allowing for the investigation of a large number of cases. In agreement with previous studies, our results revealed the lack of apoptosis in the healthy follicles (primordial, primary and secondary) when compared to the atretic ones (18, 23-25). The absence of apoptotic signals in the healthy follicles following a cryopreservation, thawing and immediate analysis lends further support to the fact that apoptosis is a dynamic process that requires active metabolism and usually takes several hours or even days to be executed (26-29). Moreover, the presence of apoptotic changes in the atretic follicles both extend the previous molecular findings (10) and lend further support to the link between apoptosis and ovarian follicular atresia. This apoptotic response in the atretic follicles may be due to the activation of endonucleases that cleave genomic DNA into oligonucleosomal fragments (30). It is also possible that substances that stimulate apoptosis may have atretogenic effects on the ovarian follicles. Of note, follicular atresia plays a critical role in recruitment of follicles for ovulation as well as in their depletion during menopause and premature ovarian failure (31). Interestingly, although many of the pyknotic nuclei and DNA globules sloughed in the cavities of the atretic follicles showed intense staining with hematoxylene, apoptosis assays revealed that they were TUNEL-negative, and . This discrepancy may be due to the inaccessibility of supercondensed DNA in these cells to the TdT enzyme. The clinical significance of our study stems from two main factors. First, the critical impact of apoptosis on the outcome of fertilization and oocyte development. In this regard, the inability to conceive is associated with a higher rate of apoptosis in their granulosa-lutein cells (32-34). Second, our study proposes the combined morphology and TUNEL assay as useful and simple strategy for the detection of apoptosis ovarian tissue destined for transplantation. Apoptosis in the cryopreserved ovarian tissues occurs with a similar localization to fresh tissue. Apoptosis in cryopreserved tissue may also occur because: during cryopreservation process, the ovarian tissues receive apoptosis-triggering signals. With a further freeze-thaw trauma; the primed cells in the cryopreserved tissues were forced to commit suicide by activation-induced apoptosis (35, 36). It is also possible that cryopreservation of these tissues was associated with degradation of apoptosis related proteins (caspase-3, caspase-8, Bcl-2 and actin) with subsequent alteration of their function and enhancement of apoptotic activity (37-39). Given the fact that the insignificantly higher apoptosis mean values were observed only in the atretic and not the healthy follicles, it is conceivable that: 1) these changes may reflect preexisting atretic changes or the occasional apoptosis in the endothelial cells and 2) cryopreservation does not seem to be responsible for any further follicular damage provided that the appropriate freeze-thaw protocol is being used. In summary, our study revealed that 1) apoptotic cell death occurs in the atretic follicles but not in the healthy ones in pig ovary; 2) combined TUNEL assay and histological examination offers a suitable method for detection of apoptosis in the ovarian tissues. It also confirms the notion that apoptosis is central to ovarian atresia. The potential value of this combined method in the clinical setting, such as in the evaluation of the integrity of the tissue for ovarian transplantation, mandates further investigations. ACKNOWLEDGEMENTS We wish to thank Dr T. Falcone MD, E Jeremias MD, A Agarwal.PhD, Department of Obstetrics and Gynecology, The Cleveland Clinic foundation, Cleveland, Ohio, USA for their kind support. REFERENCES
© Copyright 2004 - Middle East Fertility Society The following images related to this document are available:Photo images[mf04028f2.jpg] [mf04028f3.jpg] [mf04028f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}