 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
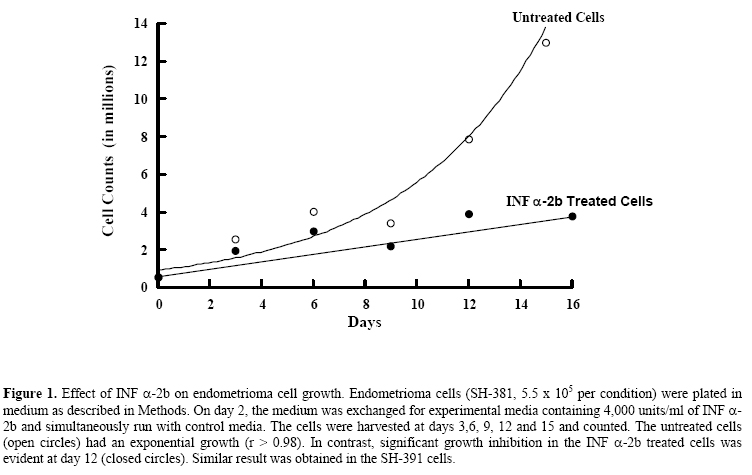
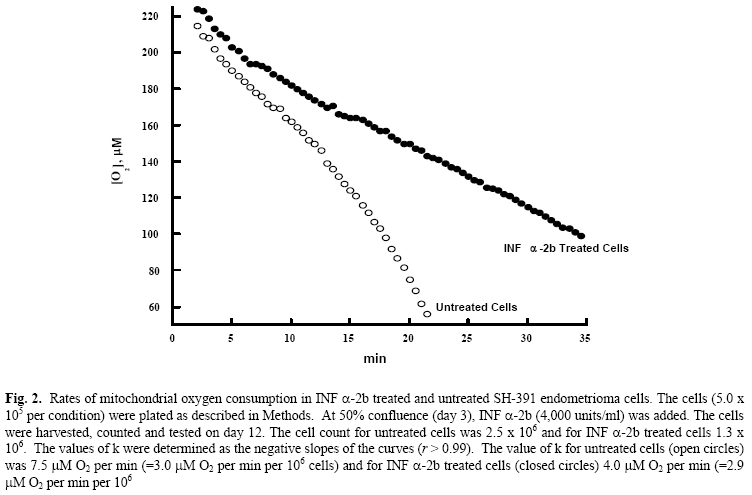
Lack of an effect of interferon α-2b on endometrioma cellular respiration Shawky Z. A. Badawy, M.D.,Violeta Cuenca, M.S.,Abdul-Kader Souid, M.D., Ph.D. Department of Obstetrics & Gynecology, Division of Reproductive Endocrinology and Infertility and Department of Pediatrics, StateUniversity of New York, UpstateMedicalUniversity, Syracuse, NY, USA Corresponding Author: Shawky Z. A. Badawy, M.D., Department of Obstetrics/Gynecology, State University of New York, Upstate Medical University, 750 East Adams Street, Syracuse, NY 13210, Phone: 315-470-7907, Fax: 315-470-2838, E-mail: badawys@upstate.edu Received on July 18, 2004; Code Number: mf04046 ABSTRACT Objective: To explore the effect of INF α-2b on oxygen consumption
of endometrioma cells in culture. Key words: Endometrioma, Interferon α-2b, O2 consumption In-vitro studies from our laboratory showed that INF α-2b invokes anti-proliferative effect on endometrioma cells (2). INF α-2b also reduces endometriosis implants in the rat model (3). Such effect is desirable for treating patients with advanced endometriosis (4). However, clinical trials using INF α-2b require further investigation of its mechanism of action in this type of tumors. It is not yet known whether INF α-2b impairs mitochondrial function of endometrioma cells. The mitochondria are a major source of cell energy. This vital organelle plays a central role in the cell integrity, and many drugs target it (5). Some agents can directly damage the mitochondria, producing rapid adenosine 5’-triphosphate depletion and non-apoptotic cell death. Others may invoke indirect mitochondrial injury, leading to loss of soluble mitochondrial proteins (e.g., cytochrome c) and activation of the apoptotic pathways (6). Identifying a drug effect on mitochondrial function is thus clinically important. The present study evaluates the effect of INF α-2b on the mitochondrial respiratory chain, using a phosphorescence analyzer that measures [O2] in solution (7-10). This method is based on oxygen quenching of the phosphorescence of metallotetra-benzoporphyrins. Our results facilitate a better understanding of the anti-proliferative activity of interferon alpha 2 beta. MATERIALS AND METHODS Chemicals INF (-2b (Intron-A) was purchased from Schering-Plough (Madrid, Spain); Pd derivative of meso-tetra-(4-sulfonatophenyl)-tetrabenzoporphyrin (Pd phosphor, sodium salt, MW~1300) was purchased from Porphyrin Products, Inc. ((Logan, UT); high flux LED based illuminator with red (peak, 630 nm) visible emission (OTL630A-5-10-66-E) was purchased from Opto Technology, Inc. (Wheeling, IL); A/D converter was purchased from Computer Boards, Inc. (Middleboro, MA); fibronectin, adenosine 5’-diphosphate (ADP), fatty acid free bovine serum albumin (BSA), ethylenediaminetetra-acetic acid (EDTA), trypsin, M199 medium, and antimycotic-antibiotic were purchased from Sigma-Aldrich (St. Louis, MO); distilled deionized water (Water For Injection quality water, dH2O) was purchased from Mediatech (Herndon, VA); phosphate-buffered saline (PBS) w/o calcium or magnesium was purchased from BioWhittaker (Walkersville, MD); fetal bovine serum (FBS) and RPMI and M199 media were purchased from Mediatech (Herndon, VA). Pd phosphor was dissolved in dH2O (2.5 mg/ml = 2.0 μM) and stored in the refrigerator for one week. Cells Endometrioma cell lines (SH-381 and SH-391) from two patients with tumors were grown in-vitro (1). Viable cells (5.5 x 105 per condition) were seeded onto T-150 flasks that were coated with fibronectin-like polymer plus. The cells were plated in M199 medium with antibiotic-antimycotic, and 10% FBS. Plated cells were incubated at 370C in 5% CO2 and given 24 hours to adhere to the flask. The use of endometrioma specimens was approved by the Institutional Review Board (State University of New York, UpstateMedicalUniversity) for the protection of human subjects. Informed consent was obtained from each patient with endometriosis. Treatment with INF (-2b) On day 2, medium was exchanged for experimental media containing 4,000 units per ml of INF (-2b, and simultaneously run with control media. The cells were harvested and counted at days 3,6, 9, 12 and 15 as described (2). Cellular respiration Cellular respiration was measured at room temperature in sealed vials containing 1-3 x106 cells per condition (7). The respiratory substrate was glucose. Cells were suspended in 0.5 ml of Pd phosphor solution [RPMI medium (containing 6.0 μM Na2HPO4 and 10 μM glucose) supplemented with 2 (M Pd phosphor and 3% BSA (final pH, 7.5)]. The solution was freshly made in a 30-ml quartz tube and continuously stirred for at least 30 min prior to use. The vials were sealed with 50 (l mineral oil and placed in the instrument. Mixing was done with a ceramic stir bar. When [O2] reached 50 (M, the cells were re-suspended in a fresh Pd phosphor solution and the measurement was repeated. Rates of cellular mitochondrial oxygen consumption were determined as the negative slopes of the curves of [O2] versus time (that is, zero-order rate constant, “k”, in (M O2 per min per 106 cells). Each experiment included a rotenone (inhibits complex I of the respiratory chain) condition. The value of k for the Pd phosphor solution without cells was (mean ( SD) 0.28 ( 0.05 (M O2 min-1, and for 107 cells incubated with rotenone (50 (M at 37°C for 1 hr) 0.36 ( 0.16 (M O2 min-1. Instrument and analysis [O2] in solution was determined as a function of time using the Pd phosphor (7-11). Its phosphorescence decay was closely characterized by a single exponential, and the decay time (t, disappearance of phosphorescence intensity) was inversely proportional to [O2]. Samples were exposed to ten light flashes per second from a pulsed light-emitting diode array with peak output at 625 nm. Emitted phosphorescent light was detected by a Hamamatsu photomultiplier tube after first passing through a wide-band interference filter centered at 800 nm. The amplified phosphorescence decay was digitized at a rate of 1 MHz by a 20 MHz A/D converter. Two hundred fifty samples were collected from each decay curve and the data from 10 consecutive decay curves were averaged for calculation of τ. [O2] was calculated using the following equation: τo/τ = 1 + τ kq [O2] (1) where τ, is the lifetime in the presence of oxygen; τo, the lifetime in the absence of oxygen; and kq, the second order oxygen quenching rate constant for the Pd phosphor in the presence of BSA (4.1 x 108 M-1 s-1) (7-10). Statistical Analysis For the cell growth experiment, data were fitted to an exponential model, yielding r value of > 0.98. For oxygen consumption measurements, linear regression was used to model the data, yielding correlation coefficient values of > 0.99. RESULTS Effect of INF α-2b on endometrioma cell proliferation Endometrioma cells (SH-381) were plated at ~5.5 x 105 cell per condition. On day 2, the medium was changed to experimental medium containing 4,000 units/ml of INF α-2b, and simultaneously run with control media. The cells were harvested and counted at days 3,6, 9, 12 and 15. The untreated cells (open circles) had an exponential growth (r > 0.98). The INF α-2b treated cells, on the other hand, had poor growth, which was more pronounced at day 12 (Figure 1). Effect of INF α-2b on endometrioma cell mitochondrial oxygen consumption The rate of cellular oxygen consumption for SH-391 cells was determined at day 12. The value of k for untreated cells (open circles) was 3.0 μM O2 per min per 106 cells and for INF α-2b treated cells (closed circles) 2.9 μM O2 per min per 106 cells. Thus, despite prolonged incubation with INF α-2b cellular respiration remained unchanged (Figure 2). DISCUSSION Interferons belong to the family of cytokines. Their discovery introduced a new aspect of our knowledge of cell biology. In addition to their immune regulatory activity, this class of drugs has anti-viral and anti-tumor activities. Interferon a is produced mainly by B-lymphocytes and macrophages. This drug was shown to reduce the size of endometriosis implants in experimental animals and human studies (2,3). It was also found to inhibit the growth of endometrioma cells in culture. Previous studies also demonstrated that these cells resume growth in culture following removal of INF α-2b, confirming preserved viability (2). Despite the well-known fact that many drugs target the mitochondria (directly or indirectly)(5,6), the effect of INF α-2b on cellular mitochondrial oxygen consumption has not been investigated. To address this question, we used a phosphorescence analyzer to explore the effect of INF α-2b on mitochondrial oxygen consumption in endometrioma cells (7-11). We conclude that INF α-2b does not produce mitochondrial dysfunction. Factors leading to the anti-proliferative effect of INF α-2b in endometriosis are not fully understood. Interferon b, for example, does not induce apoptosis. However, when interferon b was combined with the differentiating agent all-trans-retinoic acid, cell death by apoptosis was observed (12). Induction of the apoptotic genes was present only when both agents were used in combination (11). These results demonstrate that despite inhibiting cell growth, interferons do not disturb cell viability. We show here that endometrioma cells treated with INF α-2b maintain normal mitochondrial function (cellular respiration or cellular oxygen consumption). Cellular respiration was normal in endometrioma cells treated with INF α-2b. We conclude that INF α-2b halts endometrioma cellular proliferation without inhibiting their mitochondrial oxygen consumption. The results also suggest that INF α-2b is not cytotoxic. It causes regression of endometriosis by inhibiting tumor proliferation without interfering with cellular metabolism. REFERENCES
Copyright © Middle East Fertility Society |

{kind=link}
{kind=link}