 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
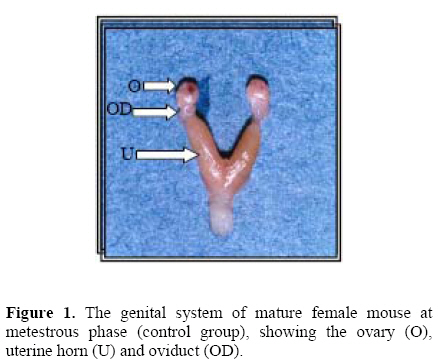
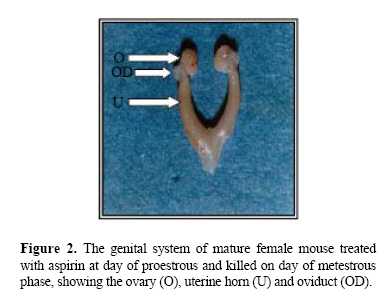
Middle East Fertility Society Journal, Vol. 12, No. 1, 2007, pp. 47-52 Pharmacological effects of low- dose of aspirin on ovulation rate in mature cycling female mice Adnan S. Al-Janabi, B.V.M.S., Ph. D.Ahmad M. Al-Zohyri, B.V.M.S., Ph. D.Fouad K. Al-Rubayai, M.B.Ch.B , M.Sc. Institute of the Embryo Research and Infertility Treatment, College of Medicine, University of Baghdad, IRAQ. Correspondence: Prof. Adnan AL- Janabi, Nahrain University, Email alnahrain_ online@yahoo.com Code Number: mf07008 ABSTRACT Objective: To determine the effects of low-dose of aspirin on ovarian
response, uterine and ovarian blood flow supply, and ovulation rate in mature
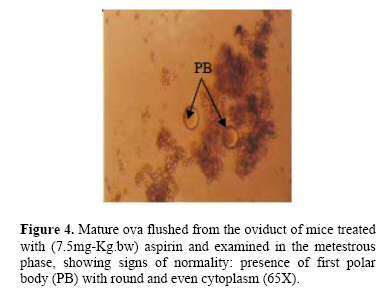
cycling female mice. Key wards: Low-dose aspirin, uterine and ovarian morphological changes, and ovulation rate. Acetylsalicylic acid was first synthesized in 1897. The product was called aspirin. In 1971, Vane described the mechanism of aspirin action, showing that it inhibits the enzyme cyclooxygenase, thus preventing prostaglandin (PG) synthesis (1). Because PGs participate in almost all human body systems, it is not surprising that, a drug with a mechanism of action related to them should have multiple therapeutic uses. Since its beginnings, aspirin has been used as an analgesic, anti-inflammatory, and an antipyretic agent. It is well known that low-dose aspirin is effective in the prevention and treatment of cardiovascular disease when given in low-doses (2–6). Furthermore, it has been demonstrated that, low-dose aspirin increased the weight of newborns in pregnant patients with fetal growth retardation, it is also used to prevent idiopathic fetal growth retardation and to improve placental and fetal blood flow in women with pre-eclampsia (7–9).Non-steroidal Anti-inflammatory Drugs (NSAIDs) acts largely through inhibition of cyclo-oxygenase (COX), an enzyme that catalyses the formation of prostaglandin from arachidonic acid (10). Aspirin, causes irreversible inactivation of cyclo-oxygenase(COX) acting on both COX-1 and COX-2 (11–13). In mice, COX-2 is rapidly induced by L.H in cumulus and granulosa cells of the ovarian follicle, with some evidence of an oocyte contribution to COX-2 expression (14). Although there are differences between primate and rodent ovaries (15), COX-2 inhibition produces the same results in both species. Prostaglandins such as PGE2 and PGF2α are critical in the process of ovulation (16), as proved by failure of ovulation in deficient phenotype mice (COX-2-1-) (17). In rats, the treatment with PG synthetase inhibitors is associated with: failure of follicular rupture; follicular hyperemia and edema (18); and suppressed synthesis of PGF2α (19). Low-dose aspirin reduced follicular level of PGF2α and PGE2, and the anti-inflammatory agent must be completely abolishes the preovulatory elevation in PG in mature follicle in order to totally inhibit ovulation in rabbits (20). In women, it has been reported that, treatment of young women with aspirin to treat inflammatory joint disease, induces luteinized unruptured follicle syndrome (LUF) (21). Moreover, aspirin or NSAIDs are associated with the recurrent development of this syndrome in young women receiving treatment with those drugs to treat ankylosing spondilytis and rheumatoid arthritis, and normal ovulation can be resumed in these patients following drug withdrawal (22). MATERIALS AND METHODS All experiments were performed on mature female Swiss white mice, 15-16 weeks old with a body weight ranging from 25-30g. The mice were obtained from the colony of the animal house of the institute for embryo research and infertility treatment, University of Baghdad. They were kept in an air-conditioned room (22-24°C) with an automatically controlled photoperiod (14 hours light and 10 hours darkness). Mice were fed the standard balanced pelleted diet presented with tap water "ad libitum". Before experimentation all mice were left for at least three weeks for adaptation. Mice then showed at least three consecutive regular cycles were included in the study (n=72 mice). Mice were then divided into treated and control groups (n=36 mice each). Treated animals were allocated into 6 equal dose group(25, 20, 15,10,7.5 and 5 mg/Kg b.w) with the aim of determining the minimal effective dose of aspirin .The preparation used was Aspegic® (Laboratories Synthelabo, France). In all cases (Treated animals=36 mice), the dose of the drug was administered twice daily, starting at the proestrous phase of the estrous cycle. The 1st drug injection was administered at 9 a.m. and the 2nd at 1 p.m. on the same day (subcutaneously). Depending on the outcome of the results of this experiment; prevention of ovulation and morphological macroscopic changes in the reproductive system, the dose level of 7.5mg/kg of b.w. was adopted as the suitable minimal dose for all other experiments in this study, (minimal inhibitory dose). Animals (36 mice) that received distilled water served as control for the experimental group. Treated mice were anaesthetized at the metestrous phase (24-84 hours after the last injection), (using Nembutal®, sodium pentoparbital - Alevetra – West Germany) and blood was collected through cardiac puncture and sera were prepared for the hormonal assay. After the killing the whole reproductive system was quickly removed after killing, and was immersed in a Petri-dish filled with in vitro medium (IVF) (Universal IVF medium, medicult, Denmark) kept at 37°C. Both ovaries were quickly dissected out, cleared from surrounding non-ovarian tissue and weighed using electronic precision balance (Sartorious – Switzerland). The uteri were then quickly dissected out slightly at the tubouterine junction from one end and immediately close to the internal orifice of the cervix from the other end, they were cleared from surrounding non-uterine tissue and were dried from IVF fluid using filter paper and they were then weighed using electronic precision balance (23). Then ova flushed from the oviduct to determine the ovulation rate. The following hormones were assayed using (Radio-immunoassay kits– RIA –), the assay was carried out in the hormonal lab. of Al-Kindy Teaching Hospital: Luteinizing hormone (LH), mIu/mL; Follicle stimulating hormone (FSH), mIu/mL. These two mentioned hormones, kits used were provided by: (cis biointernational, France). While 17β-estradiol (E2), pg/mL. Kit used was provided by: (Immunotech, Beckmann coulter company, France), by using LKB-Wallac, Minigamma 1275 Finland. The dissection of the ovaries from the oviduct was carefully performed, flushing of ova was done using tuberculin syringe (30 gauges) with blunt ended needle filled with 1mL of IVF medium was inserted into the fimbriated end of the oviduct and held in place with fine surgical forceps. Then recovered ova were kept in a Petri dish with IVF medium. And their number was counted along with their quality using a dissecting microscope under 2.5X, 16X magnification (24). Statistical Analysis Collected data were analyzed using SPSS version 10.0 for windows (SPSS, Chicago, Illinois, USA). Differences of means between groups were examined by student t-test, P. value <0.05 was considered as statistically significant. RESULTS A- Morphological changes: Aspirin administered to cycling mice on the day of proestrous and killing on the metestrous phase (i.e. during the period of ovulation), induced several morphological changes in the ovaries and uteri as follows: Ovaries appeared enlarged, cystic, congested with visually prominent blood vessels with few corpora hemorrhagica. On the other hand, the ovaries of control animals, appeared less congested, filled with corpora hemorrhagica and blood vessels on the surface of the ovaries were less prominent. Uterine changes induced congestion of the aspirin treated mice, their lumens were filled, uterine surface contain visually prominent blood vessels over both uterine horns were also seen. On the other hand, uteri of the control, appeared less congested, their lumens were also filled with fluid, but blood vessels of uterine horns were less prominent (Figures 1 and 2). B-Weight changes: Weight changes in the genital system of aspirin treated mice, showed: A highly significant (P < 0.001) increase in ovarian weight as compared to the control animals. No significant change (P=0.67) was registered in uterine weight of the treated animals as compared to the control. (Table 1) and (Figure 3). C-Number of flushed ova and their quality: Concerning effect of aspirin on ovulation rate: administration of aspirin to the cycling mice caused a highly significant (P < 0.001) decrease in the number of ova flushed from the oviduct, as compared with the control(6.00±0.37 and 11.17±0.40) for experimental and control group respectively. Administration of aspirin caused no deterioration in the quality of the flushed ova, since no ova with fragmented polar body, or degenerative oocyte, or cytoplasmic fragmentation, or vacuolated cytoplasm, or amorphic shape, or post mature oocyte were seen (Figure 4). D-Hormonal changes: No significant changes were seen in the serum level of both gonadotropins(FSH and LH) as well as estrogen(E2) in the aspirin treated mice, as compared to the control, when blood was examined on the day of metestrous phase (Table 2). Table 1.Changes in the weight of the ovaries and uteri, associated with aspirin administration to mature cycling female mice (7.5mg/Kg.bw) on day of proestrus phase and killed on day of metestrus phase.
Values are mean ± standard error (SEM), (n=6 animals/group). Table 2. Hormonal changes associated with Aspirin administration to mature cycling female mice (7.5mg/Kg.bw) on day of proestrus phase and killed on day of metestrus phase.
Values are mean ± standard error (SEM), (n=6 animals/group). DISCUSSION Ovarian and uterine morphological changes obtained in the experimental animals namely: intense organ congestion, cystic appearance of the ovaries as compared to the control are clear indication of a pronounced inflammatory response. This result is an expected outcome of a combined effect of both normal physiological morphological changes associated with ovulatory process (25), and vascular changes induced by aspirin treatment e.g.: vasodilatation and increased vascular permeability (26). All these inflammatory changes occurred normally during ovulation are thought to be mediated mainly by: prostaglandins (PGE2 and PGI2) and histamine (27, 28). Moreover PGF2α is a well known factor in terminating inflammatory changes associated with ovulation (29), and its synthesis is stimulated by gonadotropins (30) which are known to increase during this period (31). Based on these interactions we are inclined to suggest that inhibition of PGF2α by aspirin administration may have blocked or prevent the termination of the inflammatory process and allowing it to extend longer than normal. The cystic appearance of ovaries of experimental animals are due to failure of ovulation of certain number of graafian follicles which has been turned into luteinized unruptured follicles (32, 34). Inhibition of Prostaglandin synthesis in the preovulatory follicles by aspirin hinders completion of the ovulatory process leading to failure of follicular rupture a hence formation of cysts (35). Formation of these cysts has a positive impact on ovarian weight, since in ovulation, there is normally release of not only the ova but also the escape of accumulated follicular fluid, and thus decreasing ovarian weight after ovulation. These changes may explain the significant increase in ovarian weight seen in aspirin treated mice. Regarding the reduction of ovulatory rate (OR) (seen in low-dose aspirin treated group 50% decrease in comparison to the control), the underlying cause could be due to incomplete central inhibition of gonadotropin release FSH and LH which are known to be a key hormones for ovulation (36). Because Al-Bayati, (37), have shown that administration of aspirin in a very high dose (10 times a dose used in the present study) caused complete cessation of ovulation associated with significant decrease in both FSH and LH, this decrease was interpreted on the ground that it has been brought about by the suppression of PGE2 by aspirin acting centrally in the hypothalamus and locally on the ovarian prostaglandins level. Results of the present study also indicate that aspirin in the dose used was unsuccessful in inducing any significant change in the serum level of both gonadotropins. Which could mean that changes obtained (discussed above) seems to have been induced mainly on the ovarian level. Several reports have shown that, ovulation can be inhibited by NSAIDs despite of undetectable changes in several key hormones of ovulation (FSH, LH, E2, progesterone), suggesting that local ovarian factors are the predominant driving force in ovulation induction (32, 38). Many authors reported that, there is a marked increase in intrafollicular levels of prostaglandins (E and F series) shortly before ovulation (39, 40). Espey (20), reported that, low dose of aspirin partially reduced follicular level of PGE2 and PGF2α and in order to abolish ovulation totally, aspirin must be administered in high dose. Lack of any significant changes in the level of both gonadotropin (FSH and LH) in the experimental mice as compared to control, could explain absence of any significant difference in gonadal steroid in the treated and control animals REFERENCES
Copyright © Middle East Fertility Society The following images related to this document are available:Photo images[mf07008f2.jpg] [mf07008f1.jpg] [mf07008f4.jpg] [mf07008t1.jpg] [mf07008t2.jpg] [mf07008f3.jpg] | |||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}