 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
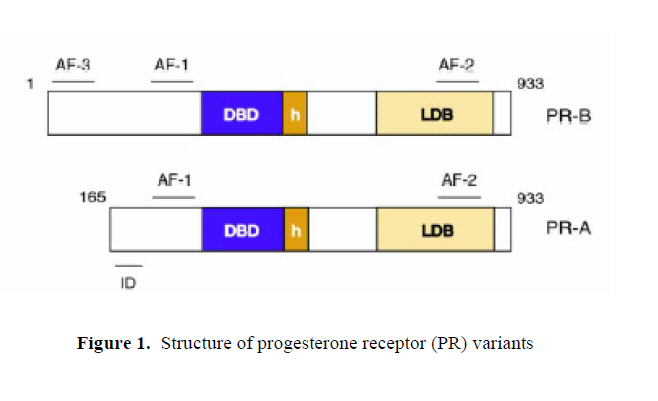
Middle East Fertility Society Journal, Vol. 12, No. 3, 2007, pp. 147-152 REVIEW Reproductive functions of progesterone Maha Al-Asmakh, M.Sc. Department of Health Science, College of Arts and Sciences, Qatar University, Doha, Qatar Correspondence: Maha Al-Asmakh, Department of Health Science, College of Arts and Sciences, Qatar University, P.O.Box 2713, Doha, Qatar e-mail: maha.alasmakh@qu.edu.qa Received on March 23, 2007; revised and accepted on April 22, 2007 ABSTRACT Progesterone has a central role in reproduction, being involved in ovulation, implantation, and pregnancy. Associated with this is the involvement of progesterone in regulation of uterine function during the menstrual cycle, by control of cyclical changes in proliferation and decidualization. Progesterone is essential for the development of decidual tissues, and if fertilization occurs, high circulating progesterone levels are important not only for facilitating implantation, but also for maintaining pregnancy by stimulating uterine growth and opposing the actions of factors involved in myometrial contraction. Key Words: progesterone, progesterone receptors, Corpus Luteum, menstrual cycle, implantation, myometrial contractility. INTRODUCTION Progesterone is essential for the regulation of normal female reproductive functions. The major physiological actions of progesterone are: a) in the uterus and ovary: induction of ovulation, facilitation of implantation, and maintenance of early pregnancy; b) in the mammary gland: lobular-alveolar development in preparation for milk secretion (1,2); c) in the brain: neurobehavioral expression associated with sexual responsiveness (3) and d) in the bone: prevention of bone loss (4). It is a 21-carbon steroid which is a precursor molecule for steroids biosynthesis. Progesterone is primarily produced by the granulosa-lutein cells of the corpus luteum (CL) during the luteal phase of the menstrual cycle and the syncytiotrophoblast of the placenta during pregnancy. However, zona fasciulata and zona reticularis of the adrenal cortex produce few amount of progesterone which is less than 1 mg/day (5). It is transported in the blood by transcortin (corticosteroid-binding globulin) and albumin with approximately 2% present in the free, unbound state. The half life of progesterone is approximately 5 minutes in the blood and its principle degradation product, pregnanediol, is formed in the liver. The plasma progesterone concentration is normally less than 5 nmol/l (1.5ng/ml) during the follicular phase of the menstrual Cycle. During the luteal phase, the plasma level rises to a peak value of approximately 40-50 nmol/l (12-16 ng/ml) (6). Action of progesterone Progesterone regulates a plethora of biologically distinct processes in a wide variety of tissues through the action of progesterone receptors. The physiological effects of progesterone are mediated by interaction of the hormone with two progesterone receptor isoforms, PR-A and PR-B. Both proteins arise from the same gene and are members of the nuclear receptor superfamily of transcription factors. PR-B differs structurally from PR-A in that it contains an additional stretch of 146 amino acids at the N-terminus of the protein. As ligand activated transcription factors, both PR-A and PR-B consist of centrally located DNA binding domain (DBD), which is flanked at the N terminus by an activation function-1 (AF-1) and at the C terminus by a hinge region as well as ligand binding domain (LBD) containing a second activation function (AF-2). PR-B consists of a third activation function (AF-3) located in the N-terminal region (Figure 1) (7). PR-B has been shown to be more potent activator of transcription of several PR targeted genes regulated by both receptors (8). Moreover, PR-A can repress the activity of PR-B (9). Function of progesterone during the menstrual cycle Progesterone is essential for the implantation and maintenance of early human pregnancy. The follicular phase of the menstrual cycle is estrogen dominated, while the luteal phase of the menstrual cycle is progesterone dominated (10). Secretion of progesterone converts an estrogen primed proliferative endometrium into a secretory one, which is receptive to the blastocyst. Before ovulation, granulosa cells in the follicle biosynthesize and secrete estrogen. After follicle rupture and release of the ovum, these granulosa cells mature to form the CL, which is responsible for secretion of progesterone and estrogen in the latter part of the cycle. In human, if fertilization does not occur within 1 to 2 days, the CL will continue to enlarge for 10–12 days followed by regression of the gland and concomitant cessation of estrogen and progesterone release. If fertilization occurs, the CL will continue to grow and function for the first 2 to 3 months of pregnancy. After this time it will slowly regress as the placenta assumes the role of hormonal biosynthesis for the maintenance of pregnancy (2). Progesterone effects on ovulation and luteinization In primates, luteinization and follicular rupture occur 36–38 h after the onset of midcycle gonadotropin surge. During this preovulatory phase, granulosa cells undergo changes in response to the ovulatory stimulus that result in terminally differentiated luteal cells. While differentiating (luteinizing) granulosa cells secrete large amounts of progesterone. The discovery that these cells express the progesterone receptor led to the hypothesis that progesterone acts in a local manner to mediate ovulation and luteinization (11). Acute administration of 3ß-hydroxysteroid dehydrogenase (3ß-HSD) inhibitors or progesterone receptor antagonists prevented ovulations in monkeys (12) and mice (13). Moreover, follicles from progesterone-depleted monkeys and progesterone receptor knockout mice (PRKO) do not luteinize (14). The increase in progesterone levels and in progesterone receptor expression within 12h of the ovulatory stimulus in the macaque follicle supports a critical early role for progesterone in ovulation and luteinization (15). However, the mechanisms whereby progesterone promotes ovulation and luteinization remain unknown. The preovulatory surge of gonadotrophins activates a cascade of proteolytic enzymes resulting in the rupture of the follicular wall and the release of a fertilizable ovum during ovulation. Several lines of evidence support a role for progesterone in the induction of proteolytic activity in the preovulatory follicle of primate and non-primate species. Levels of mRNAs for matrix metalloproteinases-1 (MMP-1) and its tissue inhibitor (TIMP-1) increased dramatically within 12h of gonadotrophin stimulus and were up-regulated by progesterone (16). Moreover, inhibition of progesterone synthesis or blocking progesterone action with RU486 decreased MMP activity in the rat and ewe (17). A regulatory role for progesterone in the activation of other ovulation-associated proteases, such as plasminogen-activator (PA), has been suggested as well, because administration of a selective progesterone receptor antagonist, Org 31710 to gonadotropin-treated rats resulted in lower PA activity levels (18). Progesterone effects on cellular proliferation in the uterus during the menstrual cycle Changes in proliferative activities of the glandular epithelium and stromal elements of the human endometrium correlate to the circulating levels of estrogens and progesterone. During estrogen-dominated follicular phase, cellular proliferations occur in both epithelial and stromal cells. This is followed by a decline in proliferation in the first half of the secretory, progesterone-dominated, phase of the cycle. In the late luteal phase, while proliferative activity remains low in the epithelium, a second peak of proliferation, consistent with decidual changes, is seen in the stromal elements. Estrogen stimulates epithelial cell proliferation, while progesterone opposes the mitotic effects of estrogen and inhibits proliferation (2, 9). In PRKO mice, ablation of both PR-A and PR-B isoforms resulted in a mark hyperplasia in the endometrial epithelium due to unopposed proliferative estrogen action (14). However, in a PR-A knock-out mice (PRAKO), in which the expression of the PR-A isoform is selectively ablated, the PR-B isoform functions to mediate rather than inhibit cellular proliferations. This gain of PR-B-dependent proliferative activity upon removal of PR-A indicates that PR-A is necessary not only to oppose estrogen-induced proliferations, but also required to inhibit proliferations induced by progesterone acting through the PR-B proteins (9, 19). Progesterone effects on cellular differentiations As a result of the inhibitory effects of progesterone on cellular proliferation, progesterone induces secretory differentiation in the glandular epithelium and stormal fibroblast. The differentiating action of progesterone is terminal: if implantation does not occur, the tissue is shed and endometrial renewal from the basal portion of the endometrium takes place. Progesterone effects on the stromal decidualization Decidualization is described as the progesterone mediated differentiation of small stromal fibroblast into large epithelioid decidual cells. This process occurs around day 23 of the menstrual cycle and is accompanied in fertile cycles by the implantation event (1). The decidual reaction is inhibited in PRAKO mice, but not PRBKO mice, suggesting a critical of PR-A in this process (9, 19, 20). In humans, decidual transformation occurs in stromal cells surrounding the spiral arties approximately 10 days after the postovulatory rise in ovarian progesterone level, indicating that the expression of the deciduas-specific gene is unlikely to be under the direct control of activated PR. Evidence has emerged to suggest that the initiation of decidual transformation requires elevated intracellular cAMP levels and sustained activation of protein kinase A (PKA) pathway (reviewed in Gellersena and Brosens, 2003) (21). Effects of progesterone on menstruation Menstruation can be defined as the shedding of the superficial layer of the endometrium due to withdrawal of progesterone following luteolysis (10). It is the result of enzymatic autodigestion and ischaemic necrosis. During the first part of the secretory phase, acid phosphatase and lytic enzymes are restricted to the lysosomes. Progesterone plays a role in stabilizing the lysosomal membranes. During the second part of the secretory phase, these lysosomal membranes are degraded resulting in the release of lytic enzymes into the cytoplasm and intracellular membrane. The lytic enzymes digest cell elements, including intracellular bridges and desmosomes (22). MMPs have an important role causing degradation of many components of the uterine extracellular matrix, including proteoglycan, glycoprotiens and basement membrane collagen (17). There is substantial evidence that MMPs are produced in the endometrium and that expression of their mRNAs is closely correlated with the process of normal menstruation (23). Several studies showed that production of endometrial MMPs is modulated by progesterone withdrawal in vitro (24) and in vivo (25). Haemostasis is the balance between coagulation and fibrinolysis. Progesterone has a role in maintaining coagulation and as a result any fall in progesterone will engender fibrinolysis and initiate menstrual bleeding (22). Vasoconstriction of the spiral arterioles also plays a role in the breakthrough of the menstrual bleeding. PGF2α causes vasoconstriction. It is negatively controlled by progesterone and causes reduction in blood flow to the corpus luteum and, thus, may cause luteolysis by depriving the gland of nutrients and substrates needed for steroidogenesis (22, 26). Finally, apoptosis takes place. Apoptosis is a phenomenon regulated by the gene bcl-2 (B cell lymphoma/leukemia-2) causing gland cell death and shedding of the menstrual blood (22). The protooncogene bcl-2 functions to prolong the survival of healthy and pathological cells by blocking apoptosis. Several studies showed a decrease in the expression of bcl-2 during menstruation and following the withdrawal of progesterone (27, 28). Effect of progesterone in the Regenerative phase Angiogenesis (new blood vessel formation) is rare in adult tissue. The female reproductive tract is an exception, with blood vessel formation taking place during regeneration, development of spiral arterioles in the late secretory phase and at the time of implantation. Three peaks of regeneration have been indicated in endometrial tissue. Two peaks of endometrial regeneration under the control of estrogen occur immediately postmenstrually and during the midproliferative phase of the cycle. The third peak is progesterone related and occurs during the secretory phase of the cycle. This peak involves the growth of spiral arterioles. The persistence of stromal progesterone receptors provides evidence that progesterone influence the development of spiral arterioles (29). Implantation Implantation of the mammalian embryo into the wall of the uterus is regulated by progesterone. Progesterone promotes implantation by stimulating production of enzymes responsible for lysis of the zona pellucida. However, while progesterone is known to be essential for implantation to occur, lysis of the zona is not the crucial step in this process, indicating that other essential progesterone-mediated events are yet to be described in the initiation of implantation (2). Implantation in PR null mice fails because the uterine stromal cells cannot undergo decidualization (14). During the time of implantation a number of progesterone dependent proteins are produced. Immunohistochemical analysis has located these proteins exclusively in the epithelial component. These progesterone dependent proteins are: progesterone –associated endometrial protein (PEP), insulin growth factor binding protein 2 (IGFBP2) or protein 14, crystalloglobulin, integrins and glycoprotiens or type 1 mucins. The physiological role of these proteins is not fully understood. Protein 14 may have immunodepressive effect thereby facilitating embryo implantation. On the other hand, type 1 mucins may inhibit implantation at the maternal cell surface (22). Uteroglobin is another progesterone regulated protein produced by the endometrial gland and apparently limited to rabbits. The function of this protein is unknown. It has been hypothesized that uteroglobin may protect the embryo from maternal immune and inflammatory response during implantation. A homolog of uteroglobin, the Clara cell 10-kDa protein (CC10), has been described in the human. However, it is expressed primarily in the lung and does not always respond to progesterone stimulation (2). Pinopodes are group of progesterone dependent proteins. The function of pinopodes is unclear; however, they may be involved in implantation by mechanisms that involve uptake of macromolecules, withdrawal of uterine fluid and facilitating adhesion of the blastocyst to the luminal epithelium (30). Myometrial contractility Progesterone prevents myometrial contractility during pregnancy. This effect is manifested by an increased resting potential and prevention of electrical coupling between myometrial cells. Furthermore, progesterone decreases uptake of extracellular calcium that is needed for myometrial contraction by downregulating expression of genes that encode subunits of voltage-dependent calcium channels. During pregnancy the adrenergic system is involved in myometrial quietening. Progesterone increases transcription of ß-adrenergic receptors in myometrium from late pregnant rats, resulting in increased sensitivity to adrenergic agents (2, 26). REFERENCES
Copyright © Middle East Fertility Society The following images related to this document are available:Photo images[mf07027f1.jpg] |
| |||||||||

{kind=link}