 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Malaysian Journal of Medical Sciences, Vol. 9, No. 1, January 2002, pp. 3-8 REVIEW ARTICLE C-Fos and Its Consequences in Pain Asma Hayati Ahmad & Zalina Ismail Department of Physiology School of Medical Sciences, Universiti
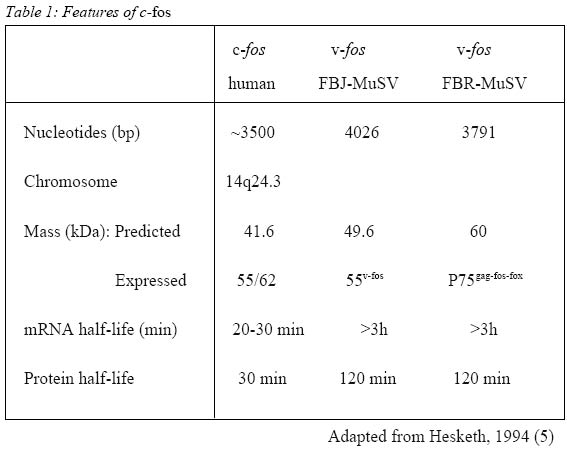
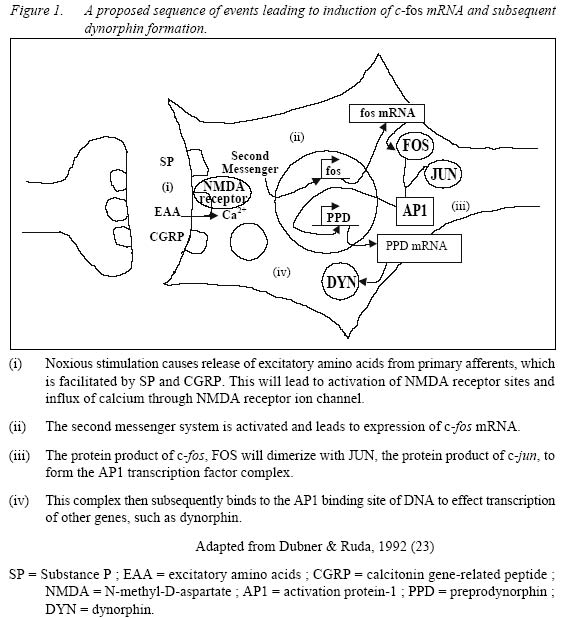
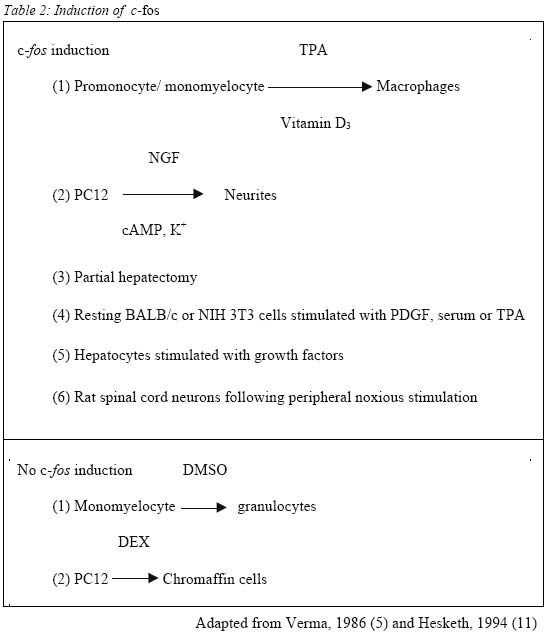
Sains Malaysia 16150 Kubang Kerian, Kelantan, Malaysia Submitted-18.9.2001 Code Numbers: mj02002 The discovery that c-fos, a proto-oncogene, has a role in pain, has triggered extensive research on the consequences of c-fos expression. It has been shown that c-fos, through its protein form, FOS, leads to expression of dynorphin gene and subsequently dynorphin protein which is implicated in the development of a pain state. This mini review looks at the properties of c-fos and the consequences of its expression following noxious (painful) stimulation. Key words : c-fos, pain, dynorphin The Human Genome The Human Genome Project, started in 1990 and completed recently, has successfully decoded the entire sequence of the human genome, i. e. the arrangement of the 2.9 billion nucleotides contained in molecules of DNA (1). There are about 30000-40000 genes in the human genome (2). Out of these, it is estimated that about 30 are known to be proto-oncogenes, and it is estimated that not more than one hundred of them occur in the human genome (3). C-fos, the cellular counterpart of the viral oncogene v-fos, is a proto-oncogene. Proto-oncogenes Ironically, the discovery of the proto-oncogene was preceded by the discovery of the viral oncogene, v-onc, which is the mutated form of the normal proto-oncogene. V-src, the first viral oncogene to be discovered, was founded by Martin in 1970 (4). Many such viruses have RNA genomes and this family of `retroviruses' replicate through a DNA intermediate in infected cells (5). Later, Stehelin et al, while screening total DNA in chickens, found that the DNA of normal chicken cells contains nucleotide sequences closely related to at least a portion of the transforming gene(s) of ASV (avian sarcoma virus) (6). These findings and further research led to the conclusion that this specific viral gene was not viral after all but derived from the normal cellular genome. These normal cellular genes are themselves highly conserved in evolution and the viral oncogenes are now recognized as sequences that are similar to normal cellular genes that have been picked up accidentally from a previous host cell (7). Thus the term c-onc and v-onc were coined, whereby the prefix v, as in v-src indicates a viral oncogene, and c, as in c-src, refers to the cellular counterpart (7). C-onc is also known as proto-oncogene, and is defined as a normal cellular gene usually associated with cell growth and cell division, and that can convert to an oncogene and promote oncogenesis. Further research led the way to the conclusion that highly conserved proto-oncogenes are present in normal cellular DNA in low copy number in virtually all members of the animal kingdom. Examples of proto-oncogenes include c-fos, c-jun, c-src, c-ras etc. c-fos The fos oncogene has been found in two retroviruses, namely Finkel-Biskis-Jinkins murine osteosarcoma viruses (FBJ-MSV) (8), and in the Finkel-Biskis-Reilly murine osteosarcoma viruses (FBR-MSV) (9,10). It is included in the group of immediate-early genes which means that its induction, i.e. conversion of the information encoded in the c-fos gene to messenger RNA, occurs rapidly (within minutes after a particular stimuli) and transiently. Its protein product, FOS, expression is maximal by around 60 minutes (11). Other features of c-fos are shown in Table 1. Among the most intriguing features of the c-fos gene are its pronounced tissue-, cell type-and stage-specific expression, its inducibility by growth factor, its differentiation-promoting potential, its regulatory mechanisms acting at the transcriptional, post-transcriptional and possibly post-translational levels, and the role of its viral homologues in malignant transformation (12). Its specificity is illustrated during normal development of the mouse whereby the highest levels of c-fos expression occur in the late gestation extraembryonal membranes surrounding the fetus (amnion and yolk sacs), in the mid-gestation fetal liver, and after birth, in whole bone marrow (13). Among the various hematopoietic cells analyzed, high levels of c-fos mRNA have been detected only in mast cells, differentiated neutrophils, blood monocytes and mature macrophages (12). All the above-mentioned tissues and cell types express c-fos at high levels in the normal environment of the organism. Other cells and tissues, in contrast, show only very low c-fos expression under the same conditions. In such cells, however, c-fos expression is inducible (12). Figure 1 shows the sequence of events leading to induction of c-fos mRNA and the formation of FOS protein. One example is in quiescent fibroblasts, where c-fos expression is normally very low, but is rapidly induced by peptide growth factors. It has also been shown that the TPA-induced macrophage-like differentiation of HL60 human promyelocytic precursor cells is accompanied by the induction of both c-fos mRNA and protein within 15 minutes after treatment (14). Despite the similarities of rapid and transient c-fos induction, there is one major distinction in the fos gene expression in resting cells and differentiating monomyelocytic cells. No c-fos transcripts are observed by 240 minutes in stimulated fibroblasts while the c-fos mRNA can be observed for 10 days during monocytic differentiation. It would thus appear that the transcriptional unit of the c-fos gene has different regulatory mechanisms in the 2 systems (14) and the c-fos gene product may serve different functions in different cell types (12). Table 2 shows other examples of cellular activation of c-fos mRNA expression in various cell types under the influence of specific agents. c-fos And Pain It was first observed in 1987 that stimulation of primary sensory neurons causes the expression of c-fos protein-like immunoreactivity in the nuclei of postsynaptic neurons of the dorsal horn of the spinal cord (15). The stimuli used to activate primary afferent inputs were noxious heat and chemical applied to the hindlimb of rats. The most extensive labeling is generated by noxious stimuli in those regions of the dorsal horn which receive small diameter primary afferents, namely Ad and C fibers. The regions involved are laminae I, II and some in lamina V. Further research showed the existence of a `second wave' of fos expression in the deep laminae (V to X), commencing at 8 hours and peaking at 16 hours; this second wave of labeling started ipsilaterally in the spinal cord and spread to become bilateral (16). On the other hand, innocuous or non-noxious stimulation such as brushing of hairs, which activates low-threshold cutaneous afferents i.e. myelinated Ab and myelinated C low-threshold mechanoreceptors and Ab/d hairs, and gentle manipulation of joints, which activates Aa proprioceptive afferents, result in fewer labeled cells with a different laminar distribution (15). No FOS expression is seen in neurons of the dorsal column nuclei, ventral horn or dorsal root ganglion cells. Although in neonatal rats c-fos can be induced in the spinal cord by both noxious and innocuous peripheral stimulation, the response to innocuous stimulation is gone by postnatal day 21 (17). This illustrates the specificity of distribution of FOS expression in response to sensory stimuli. This property of c-fos expression makes it an invaluable tool in the study of pain, as it can be used as a specific marker (18) to map out regions corresponding to noxious stimulation and the effect of analgesic or anaesthetic agents in the treatment and/or prevention of pain (19). An important role of c-fos is its implication in the development of a pain state, as part of the adaptive response of the spinal cord to continuous or subsequent nociceptive input, or both (18). In the spinal cord, following high frequency C-fiber strength electrical stimulation, there is release of excitatory amino acids aspartate and glutamate which is enhanced by substance P, glutamate and neurokinins, i.e. neurotransmitters associated with the processing of nociceptive information (20). This will result in fast synaptic potentials produced by actions at non-N-methyl-D-aspartate (NMDA) excitatory amino acid receptors, i.e. 2-amino-3-hydroxy-5-methyl-4-isoxazole-proprionic acid (AMPA) and kainic acid (KA). Continual depolarization and peptides will produce slow synaptic potentials which in turn is able to overcome the voltage-dependant Mg2+ block on the NMDA receptor. Influx of calcium through NMDA receptor ion channels and voltage-gated calcium channels occurs causing activation of a wide range of protein kinases. Nitric oxide is produced which then activates the expression of immediate early gene c-fos (21). FOS, the protein product of c-fos, interacts cooperatively with JUN, the protein product of another proto-oncogene, c-jun, to inhibit c-fos transcription (5). FOS and JUN form a heterodimer to form the activation protein-1 (AP1) transcription factor which then subsequently binds to the AP1 binding site of DNA to effect the transcription of other genes (18). The AP1 binding site is found in the genes of the opioid family such as preprodynorphin, preproenkephalin as well as nerve growth factor and a number of neuropeptides including cholecystokinin (which is known to antagonize the action of morphine and endogenous opioids) and neuropeptide Y (which is thought to have an analgesic role at the level of the spinal cord). Since the preprodynorphin gene has several AP1-like binding sites and potentially may bind FOS (as a component of the AP1-transcription complex) it suggests that FOS might lead directly to dynorphin expression. Dynorphin is an endogenous opioid acting at the kappa receptor. Studies have shown the occurrence of large increases in dynorphin gene expression and dynorphin peptide levels following inflammation and following nerve injury corresponding to increased FOS expression in the same dorsal horn cells (22, 23). Dynorphin containing neurons have direct synaptic connections with projection neurons and possibly with inhibitory local circuit neurons (23). It is postulated that these local circuit neurons are excitatory. Animal studies indicated that dynorphin is neurotoxic in the spinal cord (24). Intrathecally administered dynorphin produced moderate and very transient analgesia followed by prolonged tactile allodynia (25, 26). NMDA antagonists but not naloxone, prevented the allodynia demonstrating that the allodynia was produced by dynorphin interacting with spinal NMDA receptors and not with opioid receptors (28),. Dynorphin applied to the surface of the spinal cord induced an average 50% expansion of the receptive fields of 5 to 15 superficial dorsal horn neurons (23). Therefore it can be postulated that the large increases in dynorphin gene expression and dynorphin peptide levels following inflammation, as well as following nerve injury, are related to enhanced excitability and development of expanded receptive fields. These results show that one possible consequence of endogenous dynorphin release might be the development of chronic allodynia and hyperalgesia after a relatively minor injury. Two mechanisms were proposed for dynorphin's neurotoxic actions, i. e. ischaemia, and direct toxic effect (24). Electrophysiological studies and studies on cultured .spinal cord neurons show that dynorphin can produce neuronal damage without an ischaemic insult. Therefore, the predominant mechanism for neurotoxicity seems to be a direct interaction with the NMDA receptor complex. At least acutely, endogenous dynorphin has antinociceptive properties mediated through opioid receptors. However, its non-opioid effects i.e. neurotoxicity, hyperalgesia and allodynia, mediated by NMDA receptor, have, for the most part, overshadowed its opioid actions. From the clinical perspective, the importance of dynorphin is its potential for neurotoxicity in the spinal cord and the resultant allodynia and hyperalgesia. Thus, the prevention of dynorphin's release, by suppressing nociceptive input to the spinal cord and induction of c-fos, would be the most prudent course of action to avert a chronic pathology (24). Therefore, identifying c-fos as a marker for pain and subsequent central sensitization enables the search for drugs that are able to block its induction. CONCLUSION The importance of c-fos in the study of pain is undisputed. However, there are limitations. It is obvious that studies on c-fos is only practical in animal studies, and while the parameters in animal studies are usually very much under the control of the experimenter, in clinical studies, it is not so simple. Another limitation is that the response to pain by animals does not always imitate human responses to pain, as is the response to drugs. Having said that, animal studies nevertheless provide the basis for understanding, at molecular level, the consequences of pain and subsequently how to treat it, or rather, to prevent it. REFERENCES
Copyright 2002 - Malaysian Journal of Medical Sciences The following images related to this document are available:Photo images[mj02002t2.jpg] [mj02002t1.jpg] [mj02002f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}