 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
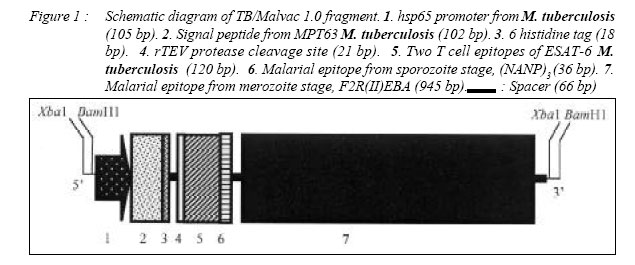
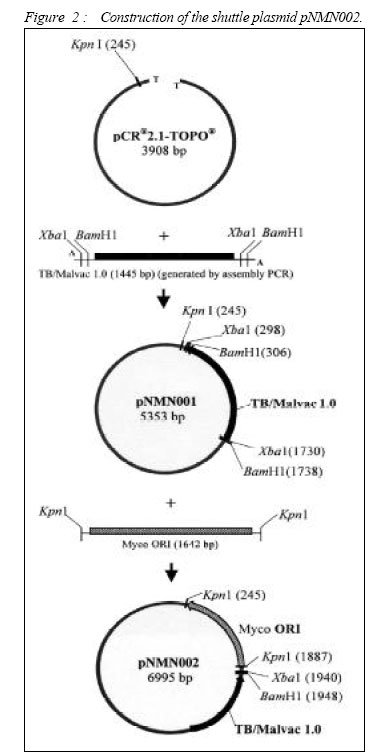
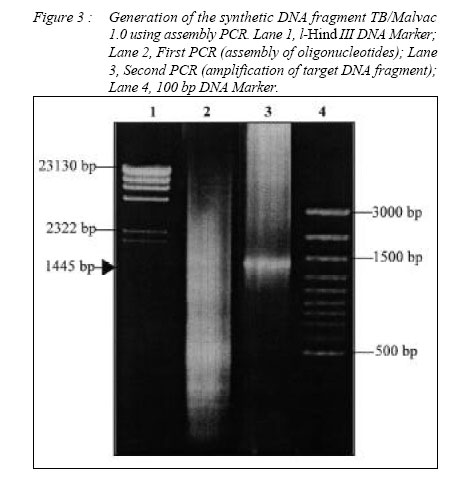
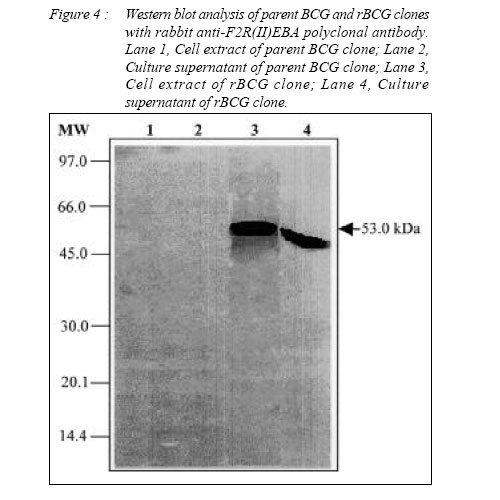
ORIGINAL ARTICLE CLONING AND EXPRESSION OF MALARIA AND TUBERCULOSIS EPITOPES IN MYCOBACTERIUM BOVIS BACILLE CALMETTE-GUÉRIN Rapeah Suppian, Zainul Fadziruddin Zainuddin, Mohd Nor Norazmi School of Health Sciences, Universiti Sains Malaysia, Health Campus 16150 Kubang Kerian, Kelantan, Malaysia Code Number: mj06003 Mycobacterium bovis bacille Calmette-Guèrin (BCG) represents one of the most promising live vectors for the delivery of foreign antigens to the immune system. A recombinant BCG containing a synthetic gene coding for the malarial epitopes namely, the fragment 2 of region II of EBA-175 (F2R(II)EBA) and the repeat sequence of the circumsporozoite protein NANP generated in favour of mycobacterium codon usage using assembly PCR was constructed. Two T-cell epitopes of the 6-kDa M. tuberculosis early–secreted antigenic target (ESAT-6) antigen were also clone in the same construct. Expression of the synthetic gene was driven by the heat shock protein 65 (hsp65) promoter from M. tuberculosis and the signal peptide from the MPT63 antigen of M. tuberculosis. Expression of the composite epitopes was detected by Western blotting of the cell extract and culture supernatant of the recombinant clones using a specific rabbit polyclonal antibody against F2R(II)EBA. This study demonstrates the possibility of cloning and expressing immunogenic epitopes from causative agents of two important diseases: malaria and tuberculosis (TB) in a single recombinant BCG construct. Key words : Assembly PCR, malaria, multivalent vaccine, recombinant BCG, tuberculosis. Introduction Malaria is one of the most serious diseases affecting much of the underdeveloped world with approximately 300-500 million people infected annually (1). Plasmodium falciparum is the most virulent of the human malaria parasites, responsible for more then 90% of malaria cases in Africa and accounts for more than one million deaths annually (2). The emergence of insecticide-resistant mosquito vectors and drug-resistant strains of plasmodium in many parts of the world has increased the intensity for the search of an effective malaria vaccine. Moreover, due to the complexity of the parasite life cycle and host immune response against malaria, it has become clear that an effective vaccine containing multiple epitopes from different stages of parasite life cycle may be required to eliminate this disease. Several candidate malarial antigens have been identified but thus far, none has shown significant and long-lasting protection in human trials (3,4). Tuberculosis (TB) is also one of the most prevalent diseases in developing countries. WHO estimates that 8.4 million new cases and approximately three million deaths occur annually (5). The only available vaccine against TB is the attenuated Mycobacterium bovis bacille Calmette- Guèrin (BCG). However, several trials conducted in different parts of the world have shown that this vaccine does not always provide consistent protection against the disease (6). Indeed, since BCG is less effective at preventing late reactivation and pulmonary TB, immunization with this vaccine has failed to control the spread of TB (7). Furthermore, multidrug-resistant TB cases have risen sharply in recent years (8,9). Thus, the development of an improved anti-TB vaccine has become an urgent necessity for adequate control and elimination of this disease. The past few years have witnessed many attempts to produce such a vaccine including one using a recombinant BCG (rBCG) containing the M. tuberculosis 30-kDa major secretory antigen (10). Despite the controversy regarding its effectiveness as a vaccine against TB, BCG has been suggested to be an attractive vehicle for the delivery of foreign antigens to the immune system (11, 12). The use of BCG as a host for the expression of foreign antigens including malaria and TB has been reported by several workers (10, 13-15). Immunization of live rBCG expressing foreign antigens has been shown to elicit good humoral as well as cell-mediated immunity directed toward heterologous antigens (14, 16-18). Although attempts to clone malarial epitopes into BCG have been reported by several workers, there seemed to be inconsistencies in the expression and immunogenicity of such rBCG clones, probably due to the difference in base composition and codon usage between plasmodium and mycobacteria (19). In an attempt to develop a multivalent vaccine against malaria and TB, we constructed a synthetic gene containing two different malarial epitopes from different stages of the P. falciparum life cycle namely the fragment 2 region II of EBA-175 (F2R(II)EBA) which is the protein that has been suggested to be involved in the sequence of events leading to erythrocyte invasion (20, 21), as well as the three repeat sequence of the circumsporozoite protein (NANP), which has been shown to elicit the production of antibodies that neutralize sporozoite activity in vitro and generates specific antisporozoite antibodies in animal models (22, 23). Two T-cell epitopes of the M. tuberculosis ESAT-6 antigen (aa 1-20 and aa 51-70), a dominant target for cell-mediated immunity in the early phases of infection in TB patients and in various animal models (24, 25) were also cloned in the same construct. In addition, we also incorporated the hsp65 promoter of M. tuberculosis and the signal peptide from MPT63 M. tuberculosis (26, 27) upstream to the epitopes. Spacer sequences were also incorporated between the epitopes in the vaccine construct to facilitate epitope-specific immune responses and to avoid antigenic competition among these epitopes. The DNA fragment was generated using a technique known as assembly PCR (28) and cloned into BCG. To increase the expression of the protein in BCG, the DNA sequence of the epitopes was optimized based on the preferred mycobacterial codon usage. Material and Methods Bacteria and mediaThe commercially available Escherichia coli TOP10 strain (Invitrogen, USA) was used in all initial cloning procedures. They were grown in LB media, supplemented with kanamycin (50 µg/ml). E. coli BL21(DE3)pLysS strain (Invitrogen, USA) was used for protein expression. They were grown in LB media, supplemented with ampicilin (50 µg/ ml) and chloramphenicol (34 µg/ml) (Sigma, USA) when required. The M. bovis BCG Pasteur vaccine strain was cultured in 7H11 or 7H9 media (Difco Laboratories, USA) supplemented with 15 µg/ml kanamycin (Sigma, USA) when required (19). Production of rabbit anti-F2R(II)EBAThe native DNA fragment of F2R(II)EBA was amplified from the cDNA of P. falciparum CAMP strain (Malaysia). The PCR product was cloned into the expression vector pRSET B (Invitrogen, USA) at the BamHI and XbaI sites to produce pNMN003 and the recombinant plasmid was transformed into E. coli TOP10. The recombinant clones were analyzed by restriction enzyme digestion and the sequence was confirmed by DNA sequencing (BioSyntech, Malaysia) before being re-transformed into E. coli BL21(DE3)pLysS for protein expression. The culture was grown overnight in LB media containing 50 µg/ml ampicilin at 37oC before being diluted to 1:10 with fresh media and further incubated until the OD600 was 0.4. Isopropylthio-βgalactoside (IPTG) (Sigma, USA) was added to a final concentration of 1 mM and the culture incubated for a further 6 hours before the cells were harvested and the proteins separated in 10% preparative SDS-PAGE under reducing conditions. The target protein was then cut out, eluted using electroelution apparatus (Bio-Rad, USA) and precipitated with four volumes of ice-cold acetone (29). Two New Zealand white rabbits were immunized subcutaneously (s.c) at multiple sites with 1 mg/ml of F2R(II)EBA fusion protein dissolved in PBS and emulsified with an equal volume of complete Fruend’s adjuvant (CFA) (Sigma, USA). The first booster was given with the same amount of protein in incomplete Freund’s adjuvant (IFA) (Sigma, USA) four weeks later, followed by a second booster eight weeks later. Blood was collected on day 0, 28, 56 and 84. The sera were separated out and stored at -20oC until used. Preparation of BCG competent cellsM. bovis BCG (Pasteur) was grown in 50 ml 7H9 media containing oleic acid/albumin/dextrose (OADC) ( Difco Laboratories, USA) for four to five days. Isoniazid (Sigma, USA) was added to a final concentration of 4 µg/ml one day prior to harvesting. The cells were harvested by centrifugation and the pellet was resuspended and washed in 25 µl prechilled 1 mM HEPES. After centrifugation the pellet was resuspended in 5 ml pre-chilled 10% v/v glycerol and centrifuged again. The cells were resuspended in 2 ml 10% glycerol and 200 ml aliquots were frozen in cold ethanol and stored at 70oC until used. Construction of the shuttle plasmid pNMN002The synthetic gene consisting of the hsp65 promoter, the signal peptide of the MPT63 protein, two T-cell epitopes of ESAT-6, (NANP)3 and F2R(II)EBA designated as TB/Malvac 1.0 (Figure 1), was generated using the preferred mycobacterium codon usage (30) by assembly PCR as previously described (19, 28). Several modifications were made to minimize duplex and hairpin formation using the OligoTm software (National Biosciences Inc., USA). Approximately one in three nucleotides was changed per codon to optimize the sequences and the G:C content were increased to approximately 60-70% in the optimized sequences. This technique involved three steps: gene assembly, gene amplification and cloning. Briefly, 73 overlapping, 24 to 46 bp oligonucleotides coding for the composite DNA fragment were mixed at 250 mM concentrations each. One ml of this mixture was then used for the first PCR for gene assembly. This was performed using the following PCR conditions: denaturation at 95oC for 1 min, followed by 55 cycles at 95oC for 1 min, 68oC for 1 min, 72oC for 1 min and a final incubation cycle at 72oC for 10 min. A second PCR was then performed using the following primers: sense 5’ ATTCTAGAGCGGATCCTGATTGAGC-CCGTCGTCGCGGG CAC 3’ and antisense 5’ CGGGATCCGCTCTAGAGGTCCTGGTCGCAGATCT 3’ to amplify the target fragment formed during the first PCR using the following conditions: denaturation at 95oC for 1 min, followed by 25 cycles at 95oC for 1 min, 68oC for 1 min, 72oC for 1 min and a final incubation cycle at 72oC for 10 min. The High Fidelity enzyme (Boehringer Mannheim, Germany) and G:C rich buffer (Maxim Biotech, USA) were used in both PCR steps. The PCR product was then cloned into the cloning vector pCR®2.1-TOPO® (Invitrogen, USA). The success of the cloning procedure were determined by restriction analysis and DNA sequencing. To clone into BCG, the plasmid was then converted into a shuttle plasmid by insertion of the mycobacterial origin of replication (Myco ORI) derived from the M. fortuitum plasmid pAL5000 (31) at the Kpn1 site. The shuttle plasmid was transformed into BCG Pasteur by electroporation. Figure 2 shows the construction of the shuttle plasmid pNMN002 for cloning into BCG. Transformation of BCGCompetent BCG cells were thawed, centrifuged and resuspended in 200 µl 10% v/v glycerol. One hundred µl of the competent cells were mixed with 100 ng DNA in chilled microcentrifuge tubes and transferred to chilled 0.2 cm electroporation cuvettes and electroporated using the Bio-Rad gene Pulser apparatus (Bio-Rad, USA) at 2.5 kV, 25 µF and 600 Ω. The cells were immediately diluted in 900 µl 7H9 containing OADC and transferred to polypropylene tubes. The tubes were incubated at 37oC for 5 hours with shaking prior to plating onto 7H11 media containing 15 µg/ml kanamycin. Expression of F2R(II)EBA by rBCGTransformants were grown to stationary phase in 7H9 media supplemented with 15 µg/ml kanamycin. Three ml of the transformants were then washed with 0.5 ml 50 mM Tris/HCl pH 7.5, resuspended in 50 µl loading buffer (50 mM Tris HCl pH 6.8, 100 mM β-mercapthethanol, 2% SDS, 10% (v/v) glycerol and 0.1% Bromophenol Blue), boiled for 5 min, and the lysate cleared by centrifugation. Protein from the culture supernatant was also harvested by precipitation with four volumes of ice-cold acetone. Ten µl of samples were electrophoresed on 10% SDS-PAGE, and the protein bands transferred to nitrocellulose filter Hybond-C extra (Amersham, USA). The membrane was blocked with 1% blocking solution (Boehringer Mannheim, Germany) and probed with the rabbit polyclonal anti-F2R(II)EBA antibody diluted 1:100 in 0.5% blocking solution. A secondary antibody, anti-rabbit IgG conjugated with peroxidase (Dako, Japan) diluted 1:1000 in 0.5% blocking solution was used and detection was carried out using the Western Blotting Detection System (Boehringer Mannheim, Germany) according to the manufacturer’s instructions. The amount of total protein loaded in the SDS-PAGE was approximately 50 µg. The initial assembly reaction involved the construction of the full-length gene from a stoichiometric mixture of the 73 oligonucleotides. This assembly PCR reaction product was then used as a template for further amplification using specific primers. Figure 3 shows the expected DNA fragment generated (1445 bp) designated as TB/Malvac 1.0. The fragment was cloned into the vector pCR®2.1-TOPO® and the success of the cloning procedure was confirmed by DNA sequencing. The plasmid was then converted into a shuttle plasmid pNMN002 by insertion of the mycobacterial replicon to facilitate cloning into BCG. The presence of the synthetic fragment TB/Malvac 1.0 was confirmed by the preparation of rBCG plasmid DNA, followed by back transformation into E. coli and restriction analysis or PCR amplification (data not shown). Expression of F2R(II)EBA by rBCGThe expression of F2R(II)EBA in rBCG was determined by Western blotting using the polyclonal rabbit anti-F2R(II)EBA under reducing conditions in the presence of β-mercapthoethanol. An immunoreactive band of approximately 53-kDa corresponding to the expected molecular mass of the TB/Malvac 1.0 protein was observed in both the cell extract and culture supernatant of the rBCG clone but not in the cell extract and culture supernatant of the parent BCG strain (Figure 4). DiscussionBCG is one of the attractive candidates for the development of live recombinant vaccines. The development of rBCG expressing protective antigens from various pathogens therefore represents a very promising possibility for efficient vaccination against viral, bacterial or parasite infection including TB (10,32) and malaria (13-16). The advantages of using BCG as a recombinant vaccine against TB and malaria is its capacity to replicate in the host macrophage and the inherent adjuvant properties of mycobacterial cell wall which may stimulate more prolonged memory T cell response (33). Indeed, BCG is safe, stable, inexpensive to mass-produce, still the most widely used live vaccine against TB and is included in the Expanded Program for Immunization of the World Health Organization as a neonatal vaccine. Therefore, the construction of rBCG expressing immunogenic epitopes of TB and malaria (the disease which vaccine is not yet available), will improve BCG vaccine efficacy against TB as well as may protect against both diseases with only a single immunization. However, the high A:T content of the P. falciparum genome may present difficulties in expression of their proteins in commonly used prokaryotic and eukaryotic expression system particularly in mycobacteria, which has high G:C content (19,34). In general, highly expressed genes are biased towards codons that are recognized by the most abundant tRNA species in the organism (35). However, we have previously shown that we could increase the expression levels of a malarial epitope in M. smegmatis if the epitope was constructed using the preferred mycobacterium codon usage (19). By employing the same technique, in this study we have demonstrated that a synthetic gene containing malarial and TB epitopes was successfully amplified and cloned into BCG. Western blot analysis using a specific antibody raised against the native version of the malarial epitope F2R(II)EBA was able to recognize the protein coded for by the synthetic gene expressed by the rBCG under the conditions used in this study. This result revealed that the composite gene fragment was successfully expressed and secreted under the control of the M. tuberculosis hsp65 promoter and the signal peptide of MPT63. The target protein of 53-kDa was detected both in the supernatant and cell extract of the recombinant clone. We have demonstrated for the first time that the immunogenic epitopes from the causative agents of two important diseases namely malaria and TB are possible to be cloned and expressed in a single rBCG construct paving the way for the development of a vaccine that will be able to protect against both diseases in the same endemic area. Immunogenicity studies to study the functionality of the vaccine construct against both malaria and tuberculosis epitopes in BALB/c mice are under way. This work was supported by the Malaysian Government IRPA Grant No. 305/PPSP/6110252. References
© Copyright 2006 - Malaysian Journal of Medical Science The following images related to this document are available:Photo images[mj06003f3.jpg] [mj06003f1.jpg] [mj06003f2.jpg] [mj06003f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}