 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
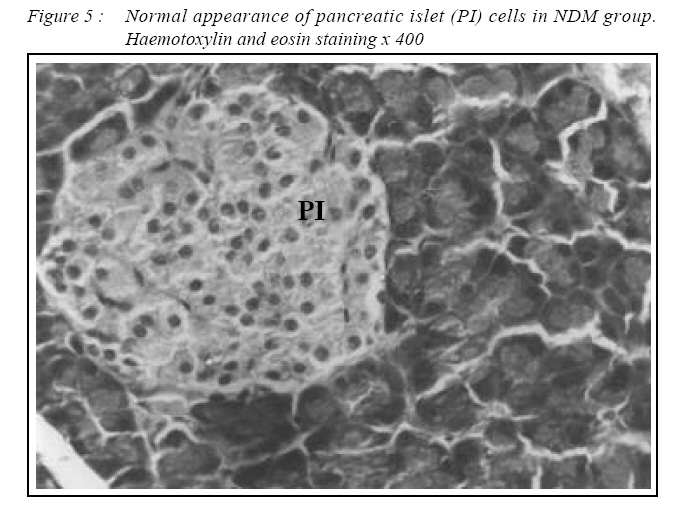
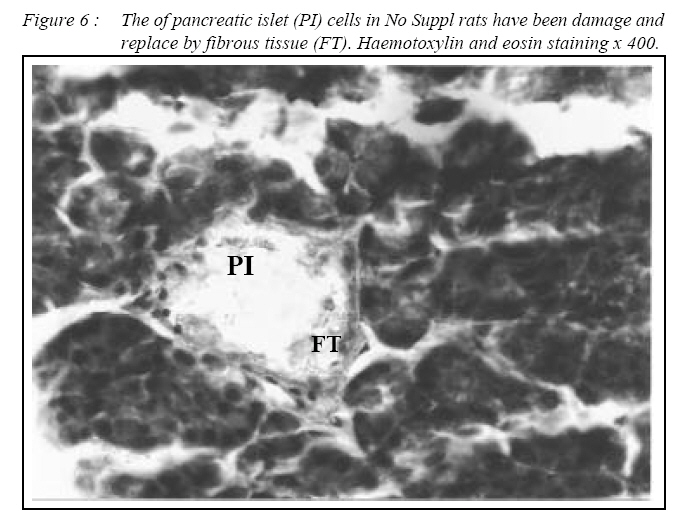
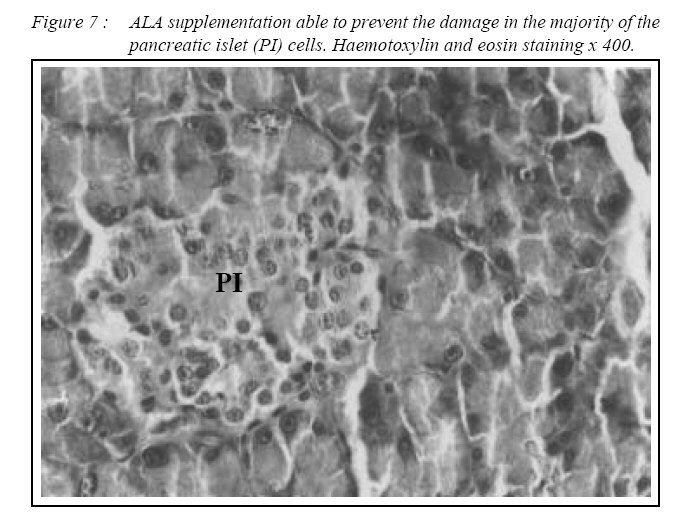
ORIGINAL ARTICLE Alpha Lipoic Acid Prevents Pancreatic Islet Cells Damage and Dyslipidemia in Streptozotocin-Induced Diabetic Rats Siti Balkis Budin, Kwa Poi Kee, Monica Yau Swee Eng, Khairul Osman, Mokhtar Abu Bakar*, Jamaludin Mohamed Biomedical Department, Faculty of Allied Health Sciences, Universiti Kebangsaan Malaysia,
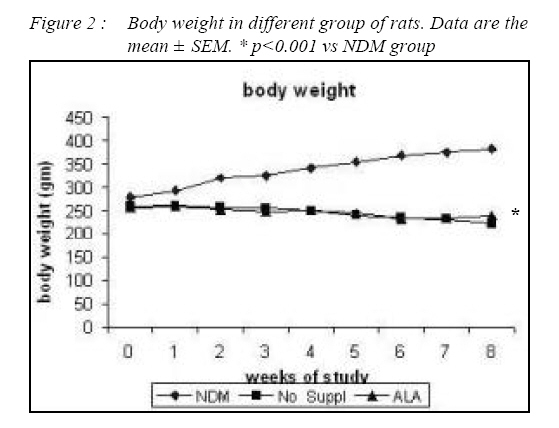
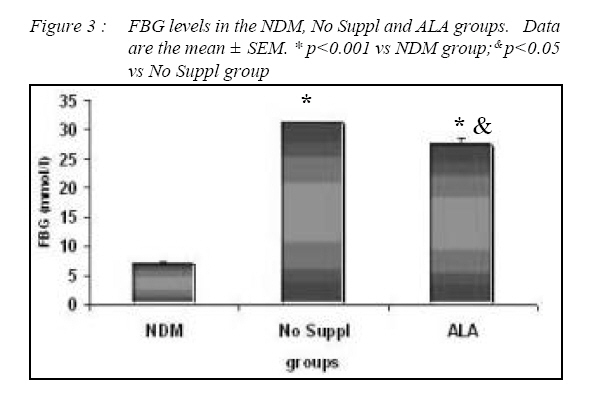
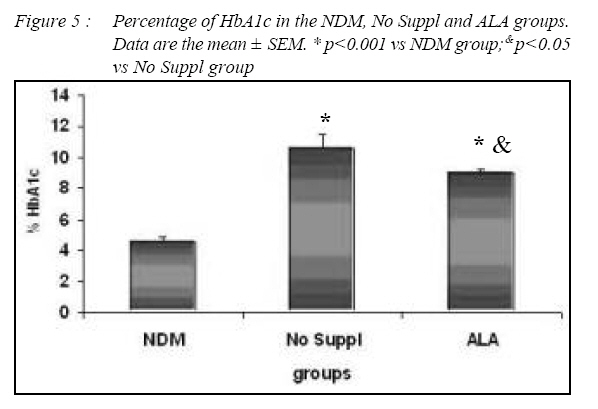
Kuala Lumpur. *Department of Pathology, Faculty of Medicine, Universiti Kebangsaan Malaysia, Cheras, Selangor Malaysia. Submitted-20-02-2005, Accepted-03-12-06 Code Number: mj07025 In the present study, the effects of alpha lipoic acid (ALA) supplementation on glycemic control and lipid profile in streptozotocin (STZ)-induced diabetic rats have been evaluated. Sprague Dawley rats were divided into nondiabetic (NDM), diabetic without supplementation (No Suppl) and diabetic with ALA groups. ALA was orally administered once a day for 8 weeks with a dose of 100 mg/kg BW. Supplementation of ALA to STZ-induced rats prevented the severe damage to the islet cells of the pancreas and lowered the plasma glucose and glycated hemoglobin (HbA1c) levels. Supplementation of ALA also suppressed the increased of total cholesterol (TC), triglycerides and low density lipoprotein-cholesterol (LDL-C) levels in the plasma of diabetic rats as well as increased high density lipoproteincholesterol (HDL-C) levels. In conclusion, this study suggest that ALA may be effective in controlling glycemic status and improving dyslipidemia in streptozotocin-induced diabetic rats and has the potential in reducing cardiovascular complications due to diabetes mellitus. Key words : Alpha lipoic acid; Diabetes mellitus; Islet cells; Hyperglycemia; Dyslipidemia Introduction Diabetes mellitus (DM) is a metabolic disease due to imbalance between glucose production and utilization (1). DM is characterized by hyperglycemia and long term complications affecting various organs such as blood vessel, nerves, eyes and kidney. Previous researchers examined oxidative stress markers in diabetic rats and found increased reactive oxygen species in pancreatic islets cells (2). In fact increased generation of reactive oxygen species such as superoxide anions, hydroxyl anions and peroxynitrite anions has been shown to occur in diabetes in association with hyperglycemia thereby deplete the activity of the antioxidative defense systems (3). Reactive oxygen species activities cause lipid peroxidation and membrane damage. Free oxygen radicals and oxidative stress appear to be an important element in the production of secondary complications in DM (3). Accumulation of lipids in DM is mediated through a variety of derangements in metabolic and regulatory processes, especially insulin deficiency. Therefore, rendering, the diabetic patient more prone to hypercholesterolemia and hypertriglyceridemia (4). One of the major pathogenesis of lipid metabolism disturbances in diabetes is the increased mobilization of fatty acids from adipose tissue and secondary elevation of free fatty acid levels in blood. Excessive lipolysis also has been found to occur in diabetes (4). As known for decades, alpha lipoic acid (ALA) is a cofactor of a-keto acid dehydrogenase complexes and plays a fundamental role in metabolism. ALA has been found to affect cellular metabolic processes, alter redox status of cells and interact with thiols and other antioxidants (5). ALA has been shown to be potentially effective in both prevention and treatment of numerous types of neurological disorders and liver diseases in which reactive oxygen species are involved (5). Although studies have provided some evidence for the beneficial role of ALA in the treatment of diabetic polyneuropathy (6), the effects of this antioxidant in carbohydrate control and lipid metabolism in experimental diabetes still remain unclear. The purpose of the present study, therefore, was to examine whether ALA has the beneficial effect on blood glucose and dyslipidemia in streptozotocin (STZ)-induced diabetic rats. Materials and Methods Experimental Design Male Sprague-Dawley rats, (each weighing 260-290g) were supplied by Animal House of Universiti Kebangsaan Malaysia, which is located at the Institute of Medical Research, Kuala Lumpur. The rats were housed in plastic containers with floors covered with wood shavings. Two rats were placed in each cage. The rats were given a standard laboratory diet and water ad libitum for eight weeks. The study was approved by Universiti Kebangsaan Malaysia Animal Ethics Committee (UKMAEC) no; Biomed/2001/Siti/17-May/047 and UKMAEC guidelines were followed. Diabetes was induced following an overnight fast, by single intravenous injection of STZ (50 mg/ kg) (Sigma, St Louis, MO, USA), which was freshly prepared in normal saline. Another group of rats received saline injection and formed the non-diabetic group (NDM) (n=7). At day three, diabetic rats with fasting blood glucose levels more than 15 mmol/l were selected for the study and divided into two groups: supplemented with ALA (n= 8) and not supplemented with ALA (No Suppl) (n=8). ALA (Sigma, St Louis, MO, USA) was administered orally at a dose of 100 mg/kg/day throughout the feeding period of eight weeks, and the supplementation was started on the same day. Food and water intake as well as the body weight were recorded weekly. Blood Analysis Following eight weeks of supplementation, rats were fasted overnight and blood was collected by cardiac puncture under deep anesthesia with diethyl ether. Blood was collected into tubes containing EDTA, kept on ice and immediately centrifuged at 3000 rpm for 20 min at 4°C. The obtained plasma was stored at - 40°C until analyzed. Fasting blood glucose (FBG) was analyzed on the same day using enzymatic glucose-oxidase kits (Trace Scientific, Melbourne, Australia, and catalogue No TR 15104). The blood HbA1c was determined using method described by Eross et al. (1984) (7) and expressed as a percentage of total hemoglobin. Determination of total cholesterol (TC), high density lipoprotein-cholesterol (HDL-C) and triglyceride levels were carried out using kits (Teco Diagnostics, 1286N. Lakeview Ave. Anaheim).Low density lipoprotein-cholesterol (LDL-C) determination was calculated using the method by Friedewald et al. (1972) (8). Statistical Analysis All results were expressed as mean ± S.E.M. The significance of the difference between the means of the different groups was obtained with one-way analysis of variance (ANOVA), followed by post hoc LSD test for multiple comparisons. The difference between groups was considered significant when p< 0.05. Results All diabetic rats showed persistent hyperphagia and polydipsia, which were not modified by ALA supplementation (Fig. 1A and B). Following eight weeks of study, there was an increased in body weight of NDM rats, whereas No Suppl rats showed a significant decrease in body weight. The decreased body weight of diabetic rats in ALA group was not modified by ALA supplementation (Fig. 2). FBG and HbA1c levels for all groups are shown in Fig. 3 and 4 respectively. FBG and HbA1c levels were found to be higher in the No Suppl and ALA groups than in the NDM group (p<0.001 for both). However, FBG and HbA1c levels in the ALA group were significantly lower (p<0.05) as compared to the No Suppl diabetic group. Fig. 5, 6 and 7 shows the histology of pancreas in NDM, No Suppl and ALA supplemented group. The histology of pancreatic islet cells was normal in NDM group. In No Suppl group, the histological section of pancreatic tissues showed severe destruction to pancreatic islet cells. Most of the endocrine cells showed degenerative and necrotic changes, and formation of fibrous tissue. On the other hand, ALA supplementation was able to prevent the destruction to occur in the pancreatic islet cells of ALA group. Levels of plasma TC, LDL-C and HDL-C and triglycerides are given in Table 1. Plasma TC, LDLC and triglyceride concentrations were markedly increased in No Suppl diabetic rats (p<0.001) as compared to NDM rats. ALA supplementation completely prevented the increase of TC, LDL-C and triglyceride and these levels were significantly lower compared with the No Suppl rats (p<0.001). On the other hand the ALA supplementation group also has significantly higher levels of HDL-C as compared with No Suppl group (p<0.001). Discussion The possible sources of oxidative stress in diabetes mellitus condition have been extensively studied for years. Diabetic patients and experimental animal models exhibit high oxidative stress due to persistent and chronic hyperglycemia (9). Under hyperglycemia, production of various reducing sugars such as glucose-6-phosphatase and fructose increases through glycolysis and the polyol pathway. During this process reactive oxygen species are produced and cause tissue damage (9). STZ is a commonly used agent in experimental diabetes. In STZ induced type 1 diabetes, hyperglycemia and oxidative stress have been implicated in the etiology and pathogenesis of disease complications (10). The mechanism by which STZ destroys β-cells of the pancreas and induces hyperglycemia is still unclear. One of the actions which have been attributed to STZ is depletion of intracellular nicotineamide dinucleotide (NAD) in islet cells. In addition, STZ has been shown to induce DNA strand breaks and methylation in the pancreatic islet cells. STZ administration also associated with the generation of reactive oxygen species causing oxidative damage to the pancreatic islet cells (11). Damaged pancreatic islet cells are clinically associated with the development of diabetes. In our study, majority of pancreatic islet cells in No Suppl rats were damaged with degenerative and necrotic changes. Pancreatic islet cells damage may lead to decrease in insulin secretion and increase in blood glucose concentration. Therefore, the blood glucose and HbA1c levels were markedly increased in No Suppl rats as compared with the NDM rats. Hyperglycemia may leads to polydipsia and polyphagia. In this study, the diabetic rats showed higher food and fluid intake as well as decreased in body weight, which also been reported by previous researchers (12). The used of antioxidant has the protective role to pancreatic islet cells against reactive oxygen species mediated damage, by enhancing antioxidants and reducing hyperglycemia in STZ-induced diabetes (13). In the present study, we examined possible usefulness of the potent antioxidant ALA. ALA prevents oxidation injury and cell death by several mechanisms, such as scavenging oxygen radicals, protecting against lipid peroxidation and chelating metal ions (6). Supplementation of ALA was shown to be capable of protecting the majority of pancreatic islet cells from damage due to STZ cytotoxicity. In the present study, ALA supplementation also reduced blood glucose and HbA1c levels in STZ-induced diabetic rats. Our results support the previous finding that antioxidant supplementation has the beneficial effects in decreasing blood glucose concentration, promoting regeneration of the pancreatic islets cells and increasing insulin release in STZ-induced diabetic rats (14). In the present study, ALA supplementation significantly reduced plasma TC, LDL-C and triglycerides and increased HDL-C in STZ-diabetic rats. The mechanism by which ALA improves the dyslipidemia is still unclear. One of the actions which might attribute to this finding is by decreasing the non-esterified fatty acid levels. Deficiency of lipoprotein lipase activity may contribute significantly to the elevation of triglycerides in diabetes. Improving the blood glucose levels served to lower plasma triglycerides levels by returning lipoprotein lipase levels to normal (15). The restoration of triglycerides levels is supported by the previous finding (15) since, ALA supplementation showed the potential in reducing blood glucose levels. The mechanism of action is also believed to be through the controlling activities of enzymes that involves in lipid metabolism. ALA was reported to reduce HMG-CoA reductase activities as well as increases the lipoprotein lipase and lecithin cholesterol acyl transferase (LCAT) activities (15). Increased LDL-C may arise from glycosylation of lysyl residues of apoprotein B as well as from decreasing affinity for the LDL receptor and hence, decreased metabolism (16). The ability of LDL-C to form lipid peroxides was found specifically responsible for the atherogenesis in diabetic patients (17). Oral administration of ALA normalized the LDL-C levels, possibly by controlling the hydrolysis of certain lipoprotein and their selective uptake and metabolism by different tissues. Since ALA also increased HDL-C, it significantly increased the anti-atherogenic index and HDL-C/TC ratio. ALA thus has the potential to prevent the formation of atherosclerosis and coronary heart disease which are secondary diabetic complications of severe DM (4). In conclusion, ALA shows the protective effects to the pancreatic islet cells and therefore improves the blood glucose levels. The alterations in lipid profiles during diabetes were also restoring to near levels by ALA supplementation. It may be concluded that the hypoglycemic effects produced by ALA may be due to the presence of antioxidant activities and the hypolipidemic effects are through the increase in lipid metabolism. But to elucidate the exact mechanism of hypoglycemic and hypolipidemic effect, further studies are essential. Acknowledgements The author is grateful for the financial support provided by IRPA Grant NO: 06-02-02-0111 and UKM grant NO: N4/2001 References
© Copyright 2007 - Malaysian Journal of Medical Science The following images related to this document are available:Photo images[mj07025f5a.jpg] [mj07025f6.jpg] [mj07025f7.jpg] [mj07025f2.jpg] [mj07025f1.jpg] [mj07025f5.jpg] [mj07025f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}