 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
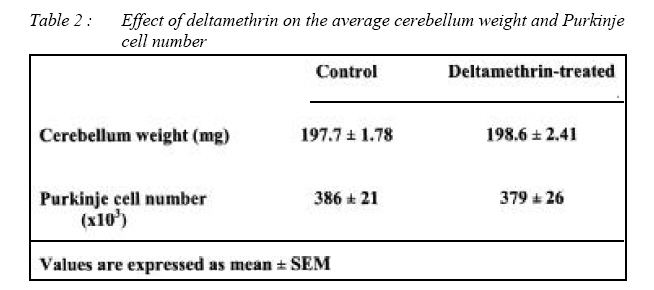
Malaysian Journal of Medical Sciences, Vol. 15, No. 3, July, 2008, pp. 14-21 ORIGINAL ARTICLE Effect of early neonatal exposure to deltamethrin on the purkinje cell number in rat cerebellum Mohd Asnizam Asari, Mohammad Shukri Abdullah, Suryati Abdullah Department of Anatomy,
School of Medical Sciences, Universiti Sains Malaysia, Health Campus,
16150 Kubang Kerian, Kelantan, Malaysia Submitted-1-6-2008, Accepted-10-10-08 Code Number: mj08022 Deltamethrin is a widely used insecticide belonging to the class of pyrethroid. Although the neurotoxicity of pyrethroids including deltamethrin is well established, it is still unclear whether exposure to deltamethrin during neonatal period has any deleterious effect on the survival of the Purkinje cells in the cerebellum. In the study, we investigated the total number of Purkinje cells in experimental rats exposed to deltamethrin using a stereological method, the fractionator. Deltamethrin in a dose of 1 mg/kg/day (corresponds to 20% of LD50 ) was administered through oral gavage to male pups from 2nd to 5th postnatal day (PND). At PND 21 the animals were sacrificed and their cerebelli were removed. The cerebelli were systematically sampled using the fractionator method and stained with cresyl fast violet. The number of the Purkinje cells was counted for each cerebellum. The results showed that there was no significant difference in the total number of Purkinje cells in the deltamethrin-treated group as compared to the control animals. This suggests that deltamethrin exposure at the current dosage during the neonatal period do not have any significant effect on the survival of the Purkinje cells in the cerebellum. Key words : pyrethroid, deltamethrin, stereology, fractionator neonatal, postnatal, nucleolus, rat Introduction Pyrethroid is considered to be among the safest class of insecticides with a relatively high insect/mammalian toxicity ratio (1). Pyrethroids are widely used in the agricultural sector to protect the crops against a wide range of insect pests. In domestic environment, they are often used as active ingredients against various household insects such as mosquitoes (2). In tropical countries, where the use of mosquito repellents is widespread, a large number of populations including the children are exposed to pyrethroids throughout the year, especially during the night (3). An investigation in Germany revealed that a large portion of the general population is homogeneously exposed to pyrethroids, most likely through the daily diet (4). Because of their widespread use, their potential health effects have become major public concern particularly in the children (5, 6). Neurotoxicity of pyrethroids on the brain in experimental animals is well established (7, 8). In general, neonatal animals are more vulnerable to the toxic effects of pyrethroids. It is estimated that neonatal rat is 17 times more vulnerable to acute toxicity of pyrethroids than the adult (9). This vulnerability is possibly due to the age-related differences in pharmacokinetics such as immaturity of hepatic metabolic detoxification (9, 10). Studies have revealed that exposure to pyrethroids during the neonatal period affected the behaviour and motor activites later in life (11, 12, 13, 14). Moreover, neonatal exposure to pyrethroids even at relatively low doses, disrupt the normal development and migration of neurons in the brain (15, 16). For instance, the administration of 0.7 mg/kg deltamethrin (< 1 % of adult median lethal dose, LD50) to the neonatal rats from postnatal day (PND) 9 to 13 was found to delay the migration of granule cells and appearance of radial glial fibers in the cerebellum. In addition, the radial glial fibers were found to be disorganized and hypertrophic after the exposure (16). Moreover, neonatal pyrethroid exposure increases the permeability of the bloodbrain barrier which may predispose the developing brain to other damaging neurotoxicants (3). Purkinje cells are located in a single layer separating the outer molecular from the inner granule cell layer of the cerebellar cortex. In rats, the Purkinje cells are formed during day 13-16 of gestation. At birth, the Purkinje cell layer is about 6-cell thick and by the PND 5, the layer reduces in thickness to form a single cell layer (17, 18). The Purkinje cells play an important role in normal development and maturation of the cerebellum and have an inductive effect on the development of the cerebellar granule cells (19). Many studies have attempted to determine the number of Purkinje cells after exposure to certain chemical substances such as alcohol (20) and nicotine (21), and after exposure to certain conditions such as x-ray irradiation (22) and malnutrition (23). A number of studies revealed that exposure to certain pyrethroids significantly affected the survival of Purkinje cells in the adult rats (24, 25, 26). While there have been a few studies on the effect of pyrethroids on the developing brain, none of the studies focused their investigation on the Purkinje cells, specifically on the number of the Purkinje cells after the exposure. Therefore, the present study aimed to investigate the effect of a type II pyrethroid, the deltamethrin, administered during neonatal period on the total number of Purkinje cells using design-based stereological method - the fractionator. Deltamethrin is a widely used pyrethroid especially in the agricultural sector and has actions against a wide range of insects (27). The fractionator is one of the design-based stereological methods to count particles such as neurons in tissue sections. It is statistically and economically more efficient than the other stereological method, so called the ‘Vref x Nv’ method in counting particles (28). The fractionator allows neurons to be estimated directly without having to make any assumptions about their size, shape or orientation. Moreover, the knowledge of organ volume is not required to calculate the total number of neurons. The major and the only requirement is that the neuron being counted can be identified unequivocally on all sections throughout the organ. The method offers another advantage, tissue shrinkage suffered during tissue processing will not interfere with the validity of the method (29, 30). Methods Animal Virgin female Sprague Dawley rats were mated and housed singly in standard plastic cages. They were maintained on standard pellet diet and water ad libitum and were checked each morning for delivery. The day of delivery was designated as PND 0. At birth, the litters were standardized to contain 8 pups with as many males as possible. The pups remained with their respective dams throughout the experimental period, except during experimental procedures. The male pups of each litter were equally divided into treated and control groups, each group consisting of 10 pups. Pups assigned to the treated group were given deltamethrin at a dose of 1 mg/kg body weight from PND 2 to PND 5 via oral gavage. The dosage of 1 mg/kg corresponds to about 20% of the median lethal dose (LD50) of deltamethrin at this age (10). Deltamethrin (99 % pure) was procured from SIGMA and prepared by dissolving it in corn oil vehicle immediately prior to the start of the experiment. Deltamethrin was administered through a polyethylene tube (1.5 mm diameter) by a hypodermic syringe. In the control group, an equal volume of corn oil was administered during the corresponding period. In both groups, the body weights were taken at PND 2, 7, 14 and 21. The present study was approved by the Animal Ethics Committee, Universiti Sains Malaysia, Health Campus, Kubang Kerian, Kelantan. Tissue Preparation At PND 21, the animals were deeply anesthetized with anesthetic ether and immediately perfused via intracardiac route with physiological saline followed by fixative. The brain was dissected out immediately after perfusion and then weighed. The cerebellum of each brain was separated from the cerebral hemispheres by cutting between the superior and inferior colliculi in the transverse fissure. The weight of the individual cerebellum was then determined. The cerebelli were kept in 10% formalin for 48 hours before they were systematically sampled with the fractionator method. Fractionator Method Estimation of Purkinje cell number in the cerebellum was determined by the fractionator method. The method utilizes a number of systematic sampling stages as described below. In stage 1 of the fractionator, each cerebellum was cut into 2 mm parasagittal slices. The position of the first slice was random in the interval of 0-2 mm. This ensures that all regions of the cerebellum had an equal chance of being sampled. All of these slices were then cut into strips of about 2 mm wide. The strips were left in the order they were cut. Every 4th strip was sampled systematically with a random start between 1 and 4. A new random number was chosen for each cerebellum. Therefore, for each cerebellum, the probability of selecting any individual strip was 1 in 4 (fl = 4). In stage 2 of the fractionator, strips sampled at stage 1 were cut into smaller pieces about 2 mm3 in size. The tissue pieces were again arranged according to the order of cutting. A systematic random sampling procedure, similar to that just described, was used to select 1 in 6 (f2 = 6) of these pieces. In stage 3, tissue sampled at stage 2 was processed for routine paraffin histology. Pieces from the same cerebellum were embedded together in the same mould. These tissue blocks were cut with a microtome (Leica) at a nominal thickness of 4 um. A sample of 1 in 30 (f3 = 30) of the sections was selected in a systematic fashion until the block had been exhaustively serially sectioned. Each selected section was picked up on glass slides and then stained with cresyl fast violet technique. To minimize experimenter biases, each of the slide was given a secret code number. The secret codes were only revealed after the completion of the counting studies. The selected slides were examined under a Nikkon (Eclipse E600) light microscope which was attached to a videocamera (CoolSnap-Pro, Media Cybernetics). Images projected from the videocamera were visualized and displayed onto a monitor screen interfaced with a computer (Dell). In the present study, the Purkinje cell nucleolus was used as the counting item. All Purkinje cell nucleoli visible in the screen were counted under the 20x objective. In the fractionator method, the particle number (ie Purkinje cell) is estimated in a known fraction of the reference space (ie the cerebellum). The total number of particles in the structure is derived by multiplying the number of objects counted in the volume sampled (ie profile of the Purkinje cells) by the inverse of the sampling fractions of the structure (29, 30). Thus the estimated total number of Purkinje cells for each cerebellum (N) is calculated by multiplying the corrected total number of Purkinje cells counted (Q) with the sampling fractions at stage 1 (f1), stage 2 (f2) and stage 3 (f3) of the fractionator. Thus, N = Q x fl x f2 x f3 = Q x 4 x 6 x 30 where Q is the corrected total number of Purkinje cell counted for each cerebellum, f1 is the stage 1 sampling fraction, f2 is the stage 2 sampling fraction, f3 is the stage 3 sampling fraction. In the present study, the total number of Purkinje cells counted were corrected by multiplying the number of counted neurons with the mean ratio of nucleolus/cell . The correction is necessary as Purkinje cells may have double nucleoli per nucleus which may lead to over counting of the cell (31). Statistics All values were expressed as mean ± standard error of mean (SEM). Statistical analysis was performed with SPSS (v12) software. Statistical significance between group means was analysed by the unpaired two-tailed Student’s t-test at p<0.05. Results Body weight Mean body weights at PND 2, 7, 14 and 21 are shown in table 1. Statistical analysis showed no significant difference in the body weights except at PND 7. At PND 7, the mean body weight of the rats treated with deltamethrin was 10.8 % lower compared to the respective control group. Cerebellum weight Mean cerebellum weights are shown in table 2. There was no significant difference in cerebellum weight between the control and treated groups (197.7 ± 1.78 vs 198.6 ± 2.41 mg). Estimated Purkinje Cell Number Estimates of Purkinje cell numbers are shown in table 2. The study found no significant differences in the estimated total number of Purkinje cells between the control (386 x 103 ± 21 x 103) and the deltamethrin-treated groups (379 x 103 ± 26 x 103). Precision The precision of the estimates can be expressed by the coefficient of error (CE) (SEM/ mean). In the present study, CE of the Purkinje cell estimate was calculated according to the method described by Gundersen et al. (32) and following the example shown by Fabricius et al. (33). The average CE for the number of Purkinje cells sampled from the control and treatment groups were 0.056 and 0.069 respectively, which were still below the recommended limit of 0.1 (34). Discussion In rats, the neonatal period is the period of critical brain development where there are active neuronal reorganization, differentiation and proliferation occurring in the brain including the cerebellum (17, 40). Drugs or toxicants which are considered relatively safe to adult may not be so during this brain growth spurt of the neonatal period. A previous study showed that exposure to certain toxicants such as alcohol during this period significantly affected the survival of neurons in certain parts of the brain and hence their number (35). Animal studies have shown that the pyrethroids have the potential to produce neuronal death in the adult. For instance, neuronal loss has been described in the cerebral cortex, hippocampal formation and cerebellum of adult rats after subchronic dermal exposure to permethrin at a dose that produced no gross clinical signs (24). Another study revealed neuronal loss in the hippocampus and cerebral cortex in adult rats after they were given a single intraperitoneal injection of deltamethrin at a dose of 12.5 mg/kg (36). In sharp contrast, several studies revealed that exposure to relatively high doses of pyrethroids did not produce significant neuronal loss in the brain. For instance, it was reported that the administration of a single dose of 20mg/kg deltamethrin (25 % of LD50) failed to induce neuronal loss in the areas of the brain examined (37). Similarly, the administration of type 1 pyrethroid, bifenthrin at doses sufficient to produce 30 hours of tremor failed to produce any detectable neuronal loss (38). In another study, the administration of near lethal dosage of type II pyrethroid, fenvalerate to rats and mice, failed to induce any pathological changes in the brain, although peripheral nerve damage was observed (39). The reasons for the discrepant nature of these findings are unknown. Currently, the data on the effect of pyrethroid exposure during neonatal period on the survival of Purkinje cells is very limited, although reasonable amounts of data exist for the adult brain. For instance, subchronic exposure to dermal alphacypermethrin at half of the LD50 caused pyknotic changes to the Purkinje cells in the adult rats (41). Another study described morphological changes in Purkinje cells after exposure to 7 mg/kg of oral deltamethrin (42). However, quantitative analysis of the Purkinje cells was not performed in both of the studies. Abdel-Rahman and his colleagues (24) attempted to quantify the Purkinje cell loss after dermal exposure to type 1 pyrethroid, permethrin, and they found a significant reduction in the density of Purkinje cells in the cerebellum after the exposure. However, unbiased stereological method was not employed in the above study. Moreover the report of density may lead to erroneous conclusion as dehydration of the specimens or sections can result in falsely higher neuronal density, so called the “reference trap”. This bias in both the control and treated groups are unknown and cannot be assumed to be the equal (31). Hence the present study employed the state of the art stereological counting technique, the fractionator, to obtain the unbiased estimate of the Purkinje cell count in the cerebellum. The present study failed to detect any significant effect of deltamethrin exposure during the neonatal period on the number of the Purkinje cells in the cerebellum. The dosage of deltamethrin used in the present study represents about 20 % of the LD50 at this age. The LD50 of oral deltamethrin in corn oil vehicle at PND 11 is 5.1 mg/kg (10). A previous study revealed that the administration of 0.7 mg/kg (i.p) deltamethrin during PND 9-13 of the neonatal period affected the cytogenesis and morphogenesis of the neurons in the cerebellar cortex. In addition, the developing vasculatures were damaged, leading to haemorrhage, thrombus formation and focal degeneration of the brain tissue (15). Another study revealed that the same amount of deltamethrin administered during PND 10-16 affected motor activity which lasted for several months (13). However, in both of the studies, no attempt was made to directly assess the effect of deltamethrin exposure on the Purkinje cells. In conclusion, our results do not provide evidence to support the notion that exposure to deltamethrin during the neonatal period causes a significant loss of Purkinje cell number in the cerebellum. Acknowledgements This work was supported by Universiti Sains Malaysia Short Term Grant (No. 304/PPSP/ 6131461). We are very grateful to Mr. Harissal for his technical support. References
© Copyright 2008 - Malaysian Journal of Medical Science The following images related to this document are available:Photo images[mj08022t2.jpg] [mj08022f2.jpg] [mj08022f1.jpg] [mj08022t1.jpg] |
| |||||||||

{kind=link}
{kind=link}