 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Medical Journal of the Islamic Republic of Iran , Vol. 18, No. 1, May, 2004, pp. 45-53 DIBUTYRYL-CAMP-MEDIATED SUPPRESSION OF DE NOVO SYNTHESIZED LIPID SECRETION IN BOTH CULTURED MCARDLE CELLS AND RAT HEPATOCYTES MEHDI RASOULI* AND RICHARD LEHNER** From the Department of Pediatrics and Cell Biology, CIHR Group on Molecular and Cell Biology of Lipids, University of Alberta, Edmonton, Canada, T6G 2S2. *Corresponding author. Assistant professor of clinical biochemistry
at University of Sari and visitor scientist at University of
Alberta. Current address: Department of Clinical Biochemistry, Medical
Sciences University of Mazandaran, Sari, Iran. E-mail address:
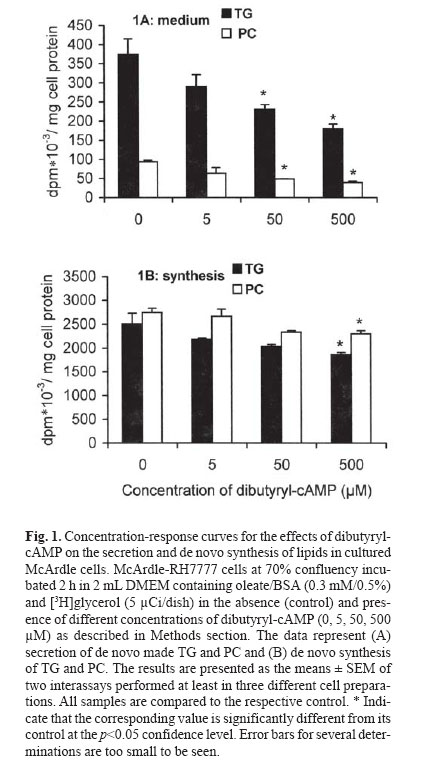
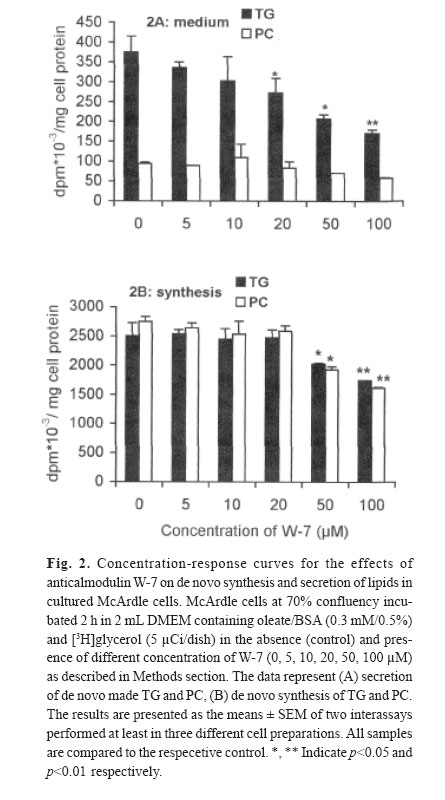
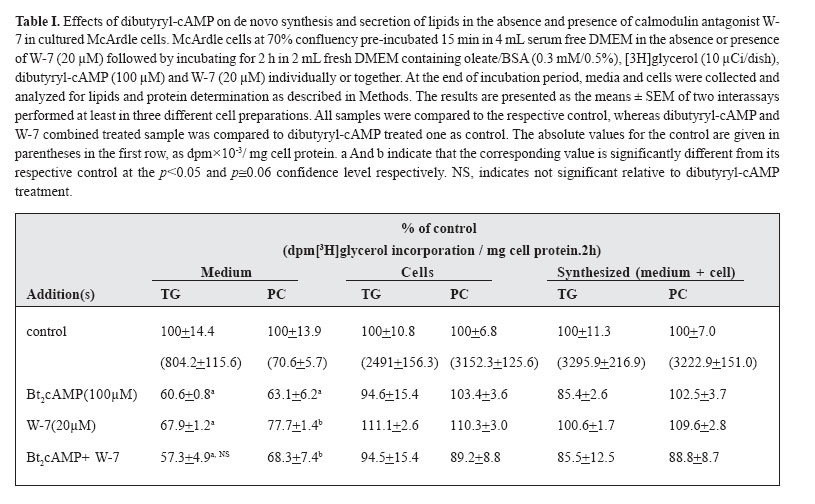
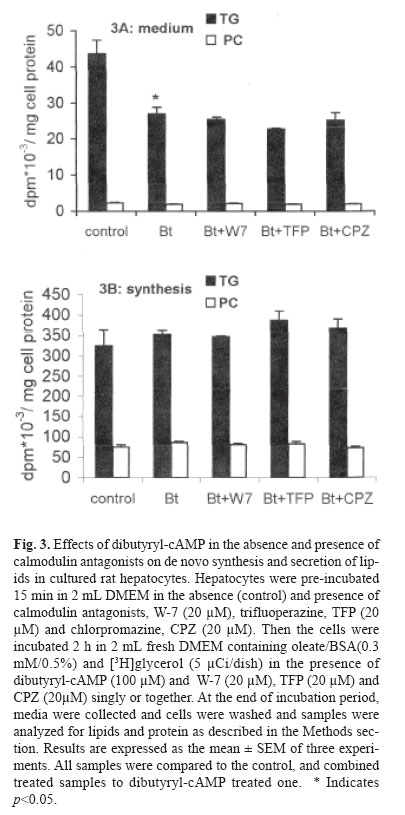
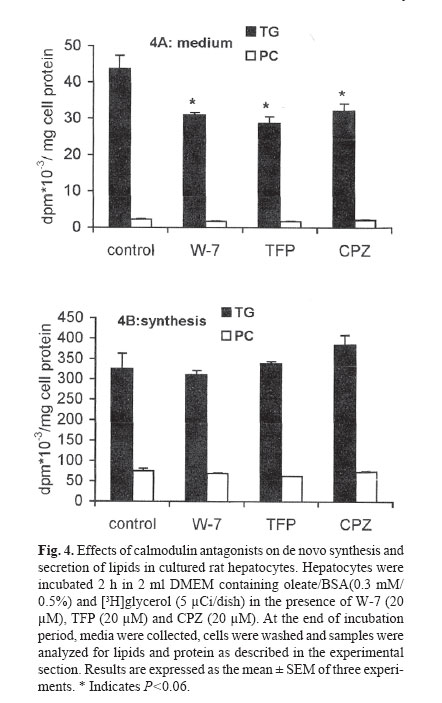
mehdi2rasouli@yahoo.com. Code Number: mr04008 ABSTRACTThe effects and interaction between cAMP-analogue dibutyryl-cAMP and calmodulin antagonists were investigated on de novo synthesis and secretion of lipids in cultures of hepatoma McArdle-RH7777 cells and normal rat hepatocytes. DibutyrylcAMP caused a significant decrease in the secretion of de novo synthesized triacyl[3H]glycerol in both cultures of McArdle cells and rat hepatocytes. The inhibitory effect of dibutyryl-cAMP was concentration-dependent and appeared at the lowest concentration examined, 5 μM. Dibutyryl-cAMP at a concentration of 50 μM suppressed secretion of triacylglycerol by approximately 38% (p<0.05) and secretion of phosphatidylcholine by 30% (p<0.05). Dibutyryl-cAMP did not affect the synthesis of triacylglycerol and phosphatidylcholine, except at the highest concentration tested, 500 μM, where both triacylglycerol and phosphatidylcholine synthesis were suppressed significantly. Anticalmodulin W-7 also inhibited secretion of newly made triacylglycerol in a concentration-dependent manner in both cultures of McArdle cells and rat hepatocytes. W-7 at a concentration of 20 μM suppressed triacylglycerol secretion by about 37% (p<0.05), while the secretion of phosphatidylcholine and synthesis of triacylglycerol and phosphatidylcholine were not affected, unless at more than 20 μM concentration, at which both triacylglycerol and phosphatidylcholine synthesis were decreased significantly. The inhibitory effect elicited by dibutyryl-cAMP (100 μM) was not abolished in the presence of calmodulin antagonists, W-7 (20 μM), trifluoperazine (20 μM) and chlorpromazine (20 μM). The simultaneous effects of dibutyryl-cAMP and either calmodulin antagonists were not additive or synergistic. None of the calmodulin antagonists affected the cellular content of de novo synthesized triacylglycerol and phosphatidylcholine significantly. Neither dibutyryl-cAMP nor any calmodulin antagonist or their combination did affect the overall rate of de novo synthesis of triacylglycerol and phosphatidylcholine. All calmodulin antagonists examined alone also had a net significant inhibitory effect on the secretion of newly made triacylglycerol. The results presented here suggest that calmodulin antagonists have net direct effects and hence could not overcome dibutyryl-cAMP-induced suppressive effects on the secretion of newly made triacylglycerol. The cell types, normal hepatocytes relative to hepatomas, did not influence the results. Keywords: Calmodulin, Cyclic AMP, Hepatocyte, Phosphatidylcholine, McArdle cells, Triacylglycerol and VLDL. Abbreviations used: Apo; apolipoprotein, BSA; bovine serum albumin, Bt2cAMP; dibutyryl cyclic adenosyl monophosphate, CE; cholesteryl ester, DMEM; Dulbecco’s modified Eagle’s medium, ER; endoplasmic reticulum, PC; phosphatidylcholine, TG; triacylglycerol, VLDL; very low density lipoprotein. INTRODUCTION The intrahepatic metabolism of VLDL is subject to control by both cAMP and calcium/calmodulin signal transduction pathways. The secretion of VLDL has been reported to be suppressed not only by calcium-linked agents such as catecholamines,1-3 prostaglandins,4 and calcium antagonists5-6 but also by agents acting via cyclic AMP (cAMP) pathway including glucagon,7-9 cAMP dependent protein kinase9 and cAMP derivatives.9-11 There are several interactions between these two transduction pathways that may lead to the regulation of apoB containing lipoproteins metabolism. There is some evidence that the effect of cAMP is mediated at least partially, via the Ca2+/calmodulin pathway14-19 Stimulation of cAMP pathway raises levels of inositol-1, 4, 5 triphosphate (IP3) and mobilizes Ca2+ in rat and guinea-pig hepatocytes12,13 and bovine parotid acinar cells.14 The cAMP analogues and forskolin (an adeylate cyclase activator) also raised cytosolic calcium in rat hepatocytes.15 Cyclic AMP also induces “capacitative calcium entry” in rat hepatocytes.15 It is reported that, calmodulin antagonist N- (6-aminohexyl)-5-chloro-1-naphtalene sulfonamide (W-7) abolished cAMP-mediated effect of calcitonin on VLDL secretion in rat hepatocytes.16 W-7 also antagonized ACTH or cAMP-induced cholesteryl esters hydrolysis and steroidogenesis in adrenocortical cells.17,18 However in order to examine the role of the Ca2+/calmodulin pathway on cAMP mediated events, it is required that the anticalmodulin drugs themselves do not exert any direct side effects. W-7 and anti-psychotic drugs such as phenothiazines trifluoperazine and chlorpromazine have been known to function as calmodulin antagonists.20 In the present investigation, the hypothesis that the effect of cAMP is mediated via Ca2+/calmodulin was examined in cultured rat hepatoma McArdle-RH7777 cells and primary rat hepatocytes. The results demonstrated that calmodulin antagonists have a net direct suppressive effect on the secretion of lipids in both cell types, and hence, they cannot overcome the effects of dibutyryl-cAMP. MATERIAL AND METHODS Materials Dibutyryl-cAMP, BSA (essentially fatty acid-free), collagenase 330 U/mg and W-7 (N-(6-aminohexyl)-5- chloro-1-naphtalene sulfonamide), trifluoperazine and cholorpromazine were purchased from Sigma (St. Louis, MO). [1(3)-3H]glycerol (2.6 Ci/mmol) was obtained from Amersham Canada (Oakville, Ontario, Canada). Dulbecco’s modified Eagle’s medium (DMEM), sodium pyruvate, penicillin/streptomycin, and fetal bovine and horse sera were from Gibco BRL (Life Technologies Inc., Grand Island, NY). TLC plates were purchased from Mandel Ltd. (Canada). All other chemicals and solvents were of reagent or better quality and were obtained from local suppliers. Growth of McArdle cells Wild typed McArdle RH-7777 cells obtained from ATCC were cultured in 60 mm dishes with DMEM containing pyruvate, antibiotics, 10% (v/v) fetal bovine serum and 10% (v/v) horse serum. When the cells reached about 70% confluency, the medium was changed to serum- free DMEM experimental medium as described in the legends of figures. All cultures were maintained in 100 mm dishes (Corning) at 37C° in humidified air (90% saturation) containing 5% CO2.21 Hepatocytes preparation, culture and incubationL6 -hepatoctes were isolated under sterile conditions from male Spargue-Dawley rats (150-200 g) by a two-step collagenase method. In brief, the liver was perfused in situ with Ca2+ free Hank's saline buffer containing EGTA (0.5 mM) for 1 min., followed by Hank's solution containing Ca2+(2 mM) and collagenase (200 IU/ml) for 9 min.21 Isolated hepatocytes were suspended in DMEM containing 15% fetal calf serum and centrifuged two times at 50 g for 2 min. The final cell suspension was counted in the presence of 0.04% trypan blue. Exclusion of the dye was >90% in all preparations. Cells were plated in DMEM containing 15% calf serum in collagen-coated (87 μg/mL) 60 mm dishes for 6h in an atmosphere of air/ CO2 (19:1). Afterwards, medium and non-adherent cells were discarded and adherent cells washed with 3×2 mL of fresh DMEM. Cells were incubated in 2 mL of DMEM containing oleate/BSA (0.3 mM/0.5%, ν=5) and other drugs as described in the legends of figures. At the end of the incubation period, the cells were cooled to 4C° on ice and the medium was collected. The cells were washed with ice cooled phosphate-buffered saline (PBS) and removed from the dish as described previously.21 Cells and media were analyzed for assays of lipids and protein. Lipid analysisAt the end of incubations, media were separated from the cells and cells were washed with ice-cold phosphate-buffered saline, harvested in the same buffer and dispersed by brief sonication. Cellular and media lipids were extracted according to Folch et al. in the presence of non-labeled lipid carrier.22 The lipids were applied to thin-layer chromatography plates and developed to 1/3 the height with chlo-roform/methanol/acetic acid/water (25:15:4:2 by vol.) to separate glycerophospholipids, followed by development in hep-tane/isopropyl ether/acetic acid (60:40:4 by vol.) to separate neutral lipids.21,23 The lipid spots were visualized by exposure to iodine, and bands corresponding to various lipid classes were scraped and the associated radioactivity was determined by scintillation counting. Other methodsProtein concentration was determined by the method of Bradford by the Bio-Rad protein assay kit using BSA as a protein standard. Oleate/BSA complex was prepared as described previously.23 The final concentration of ole-ate/BSA was 0.3 mM/0.5% (molar ratio of oleate to BSA is ν=5). Statistical analysisThe significant differences between samples and corresponding control were accessed by t-student test. The results are presented as the means ± SEM of two inter-assays performed at least in three different cell preparations. RESULTS The effects of increasing concentration of dibutyrylcAMP on de novo synthesis and secretion of lipids in cultured McArdle cellsIt has been reported that the cAMP transduction pathway is attenuated in hepatoma cells relative to normal hepatocytes,24,25 hence concentration dependence of dibutyryl-cAMP was analyzed on de novo synthesis and secretion of lipids in cultured McArdle cells. Dibutyryl-cAMP suppressed secretion of de novo made TG in a concentration-dependent manner (Fig.1A). The concentration of dibutyryl-cAMP required for significant inhibition of TG secretion was 50 μM. At this concentration, the suppression of TG secretion was approximately 38% (p<0.05). The secretion of PC was also decreased by 30% (p<0.05). Dibutyryl-cAMP had no significant effect on the cellular content of [3H]glycerol-labeled TG and PC, except at the highest concentration examined, 500 μM (data not shown). The overall rate of de novo synthesis of TG and PC was calculated from the sum of the labeled medium and cellular TG and PC when cells were cultured in the presence of exogenous [3H]glycerol (Fig.1B). Dibutyryl-cAMP had no significant effect on de novo synthesis of TG and PC, except at the highest concentration used (500 μM), where both TG and PC synthesis were suppressed 26% (p<0.05) and 16% (p<0.004) respectively. The effects of increasing concentration of W-7 on de novo synthesis and secretion of lipids in cultured McArdle cellsIt has been reported that the calcium/calmodulin transduction pathway is more sensitive to stimulation in hepatoma cells compared to normal hepatocytes.24,25 Hence, concentration-dependence of the effect of W-7 was examined on de novo synthesis and secretion of lipids in cultured McArdle cells. W-7 suppressed secretion of newly made TG in a concentration-dependent way. W-7 at 20 μM inhibited secretion of TG by about 37% (p<0.05). W-7 had no significant effect on PC secretion unless at concentrations higher than 20 μM, where PC synthesis is inhibited significantly. W-7 had no significant effect on the cellular content of [3 H]glycerol-labeled TG and PC (result not shown) and on de novo synthesis of TG and PC (Fig.2B), except at more than 20 μM both TG and PC synthesis were suppressed by 20% (p<0.05) and 30% (p<0.01) respectively. Effects of W-7 on dibutyryl-cAMP-induced suppression of the secretion of de novo synthesized lipids in cultured McArdle cellsIn order to investigate the possible involvement of calcium/calmodulin in the secretion of lipids from McArdle cells, the effects of dibutyryl-cAMP and W-7 were examined individually or together (Table I). The concentration of dibutyryl-cAMP (100 μM) and W-7 (20 μM) were chosen since at these concentrations the agents did not have any significant effects on de novo synthesis of glycerolipids TG and PC. In addition, 100 μM concentration of dibutyryl-cAMP is near the concentration for half of the maximum effect (EC50). Dibutyryl-cAMP inhibited the secretion of de novo made TG by 39% (p<0.05) and also PC by 37% (p<0.06). Since W-7 itself caused a net decrease in TG secretion by 38% (p<0.05), it could not antagonize the inhibitory effect of dibutyryl-cAMP. The simultaneous effect of dibutyrylcAMP and W-7 was not additive or synergistic. Neither dibutyryl-cAMP nor W-7 had any significant effect on de novo synthesis of TG and PC. Calmodulin antagonists did not reverse the inhibitory effect of dibutyryl-cAMP on the secretion of lipids in cultured primary rat hepatocytesIn order to clarify further the inhibition of lipid secretion by calmodulin antagonists, primary rat hepatocytes which are a more physiological model for lipid metabolism were treated. As in the hepatoma cells, dibutyrylcAMP at concentration of 100 μM inhibited secretion of TG 38% (p<0.05) from primary rat hepatocytes, while PC secretion did not change significantly (Fig.3A). Dibutyryl-cAMP did not appear to have any significant effect on de novo synthesis of TG and PC. All calmodulin antagonists examined W-7 (20 μM), trifluoperazine (20 μM) and chlorpromazine (20 μM) could not overcome dibutyryl-cAMP mediated suppression of the TG secretion (Fig.3A). Other concentrations of calmodulin antagonists were tested, however, none of the calmodulin antagonists at any concentration could reverse dibutyryl-cAMP-mediated effects (results not shown). The inhibitory effect of dibutyryl-cAMP and each of calmodulin antagonists were not additive or synergistic. None of the individual or combined treatments had any inhibitory effect on TG or PC synthesis (Fig.3B). The effects of calmodulin antagonists on de novo synthesis and secretion of lipids in cultured rat hepatocytesThe secretion of newly formed [3 H]glycerol-labeled triacylglycerol was suppressed in the presence of W-7 by 29% (p<0.06), trifluoperazine 34% (p<0.06) and chlorpromazine 26% (p<0.06) (Fig.4A). The secretion of newly synthesized phosphatidylcholine was also inhibited concomitantly with triacylglycerol but not significantly. All calmodulin antagonists examined here had no significant effect on triacylglycerol and phosphatidylcholine synthesis at least at the concentration employed (Fig.4B). Thus it seems that suppression of TG secretion at the concentration of calmodulin antagonists <20 μM is not related to the inhibition of glycerolipids TG and PC synthesis. DISCUSSIONThe data presented here revealed that, cAMP analogue dibutyryl-cAMP inhibits TG secretion in hepatoma McArdle cells at the same concentration observed for normal rat hepatocytes6-9 and perfused rat liver.10 It has been reported that, cAMP transduction pathway is modified in hepatoma cells.24,25 The low level of cAMP in hepatoma cell lines is related to less adenylate cyclase and more phosphodiesterase activity.13 However none is eligible to dibutyryl-cAMP, since it is not produced by adenylate cyclase and is not susceptible to hydrolysis by phosphodiesterase.10 Bjornsson et al. have shown that activation of cAMP pathway via protein kinase-A inhibits the secretion of apoB containing lipoproteins in a situation that TG and PC synthesis were unaffected.9 It has also been reported that, dibutyryl-cAMP at low concentration did not affect cholesterol biosynthesis.9,11 Hence, the inhibitory effect of cAMP at low concentration on TG secretion is not attributed to the inhibition of TG, PC and cholesterol synthesis. There is a dual reciprocal control over the esterification and oxidation of fatty acids.9 Inhibition of de novo synthesis of fatty acids accompanied by channeling of exogenous fatty acids and fatty acids released by lipolysis of endogenous TG stores to oxidative pathway would be a probable mechanism via which dibutyryl-cAMP diminished TG secretion. Secretion of VLDL-associated components is down regulated by both signal transduction pathways; cAMP and calcium/calmodulin. It have been shown that both α-adrenoceptor agonist phenylephrine and β-adrenoceptor agonist isoproterenol suppress secretion of triacylglycerol in freshly prepared hepatocytes.3 The effects of glucagon and isoproterenol are also exerted via two signal trunsduction pathways.12,26,27 However by now, the partition and interaction between these two pathways on the regulation of VLDL secretion have not been determined. According to "dual signaling" hypothesis there is much inter- and intra-cross-talk between the two transduction pathways.15 At the present time, the mechanism(s) whereby cAMP evokes cytosolic calcium is not established accurately, however, it has been shown that cAMP via protein kinase-A (PKA) phosphorylate Gq, IP3 -receptor and calcium channels.15 On the basis of the several reports12-18 this idea arose that the effects of cAMP, at least partially may be mediated via Ca2+/calmodulin pathway. To investigate the interaction between cAMP and calcium/calmodulin pathways, it is required that calmodulin antagonists alone have no net direct effects. Our results have shown that anticalmodulins due to the net side effects cannot overcome the inhibitory effects elicited by cAMP-analogue on triacylglycerol secretion. In the present study, anticalmodulins unexpectedly inhibited the secretion of de novo made TG in both cultured hepatoma McArdle cells and normal rat hepatocytes. This finding about the net effects of calmodulin antagonists is in agreement to some reports20, 28-33 and opposite to others.3,16-18,34 W-7 at all concentrations examined here, suppressed the secretion of lipids, however different mechanism(s) are probably involved at various concentrations. At W-7 concentration equal or less than 20 μM the effect cannot be attributed to the rates of glycerolipids synthesis since TG and PC synthesis were unaffected (Fig.2B). The first explanation for the net inhibitory effect of W-7 on the secretion of lipids in hepatoma McArdle cells may be related to the nature of the cells. It is reported that the pathways of calcium/calmodulin and cAMP have been modified in hepatoma cells relative to normal rat hepatocytes.24,25 In hepatoma cell lines, the calcium/calmodulin pathway is more, while the cAMP-signaling pathway is less sensitive to stimulation. Hence, we have done similar experiments on both cell lines to exclude the possibility of the response to anticalmodulins due to the modified nature of hepatoma cells.35 However, similar results were obtained on both cell lines. Therefore, the assumption that, the response of McArdle cells to calmodulin antagonists is related to the modified nature of hepatoma cells will be ruled out. The effects of anticalmodulins may be related to inhibition of calmodulin.6 Calmodulin activity may be involved in assembly and secretion of apoB containing lipoproteins4 and they probably would antagonize calmodulin- mediated effects that lead to suppression of lipids secretion. Available knowledge about the role of Ca2+/calmodulin in the regulation of hepatic VLDL metabolism is limited. Intraluminal calcium of ER is required for proper folding and translocation of nascent apoB through secretory pathway.36 Calcium mobilizing agents release Ca2+ from intracellular stores (predominantly ER) to cytosol. Depletion of intraluminal Ca2+-stores prevents secretion of apoB associated components,4 at the same time the increment in the cytosolic Ca2+ leads to activation of the calcium/calmodulin pathway20 and also trigger the microtubule-dependent exocytosis.37 Increment of cytosolic calcium leads to activation of exocytosis in all cells except in hepatocytes that lead to inhibition of VLDL secretion.2 Other mechanisms also may be involved. Anticalmodulin drugs have side effects that are related to nonspecific binding to other calcium-binding proteins (enzymes, receptors) and different membranes (plasma and ER membranes).20 Calmodulin antagonists bind to hydrophobic domain at calmodulin; the selectivity of binding was abolished at 10 μM or greater concentration of antagonist, since antagonists bind to other cal-cium-binding proteins.20 Unfortunately, by now it has not been possible to choose the appropriate concentration specific for only anticalmodulin activity. Indeed, many actions of calmodulin antagonists on membrane structure and hormone receptors occur at much lower concentration than needed to block calmodulin.20 It has been reported that, W-7 at low concentrations that we used here, enters into cultured fibroblasts and HepG2 cells and via interaction with In conclusion, the inhibitory effects of cAMP on VLDL secretion were not reversed by anticalmodulins. In fact, calmodulin antagonists presented at concentrations that did not alter overall glycerolipid biosynthesis also inhibited TG and PC secretion from both rat hepatoma and primary rat hepatocyte cells. ACKNOWLEDGEMENTWe wish to thank Suzzane Lingrell for preparations of primary rat hepatocyte cultures and Dr. David N. Brindley for discussions. This work was supported by the grants from the Heart & Stroke Foundation of Alberta, NWT & Nunavut and the Alberta Heritage Foundation for Medical Research. Dr. Lehner is Alberta Heritage Foundation for Medical Research Scholar. REFERENCES
Copyright 2004 -Medical Journal of the Islamic Republic of Iran The following images related to this document are available:Photo images[mr04008f4.jpg] [mr04008t1.jpg] [mr04008f1.jpg] [mr04008f3.jpg] [mr04008f2.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}