 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Medical Journal of the Islamic Republic of Iran , Vol. 18, No. 1, May, 2004, pp. 55-60 THE CYTOTOXICITY PATHWAY OF NATURAL KILLER CELLS IN CORD BLOOD COMPARED TO PERIPHERAL BLOOD M. KHEIRANDISH,*** M. EBTEKAR,* A.A. POURFATHOLLAH,*** Z.M. HASSAN,* AND A. KAZEM NEJAD* From the *Department of Immunology, School of Medical Sciences, Tarbiat Modarres University, Tehran, and the **Iranian Blood Transfusion Research Center, Tehran, Iran. Email: kheira_m2001@yahoo.co.uk Code Number: mr04009 ABSTRACT
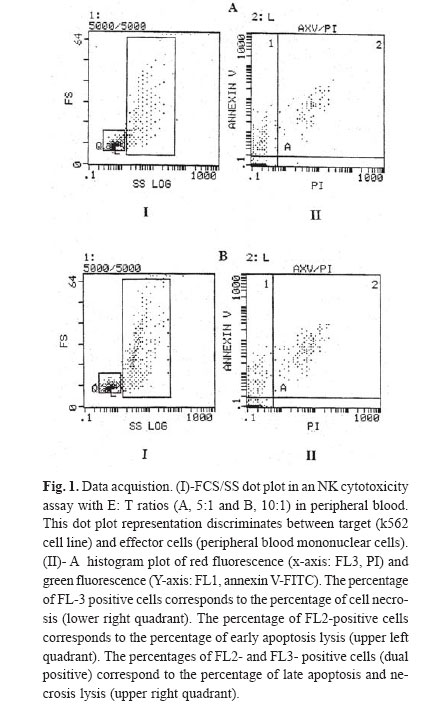
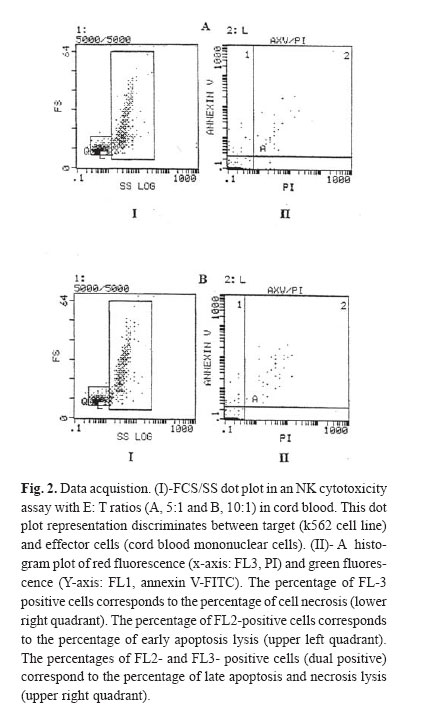
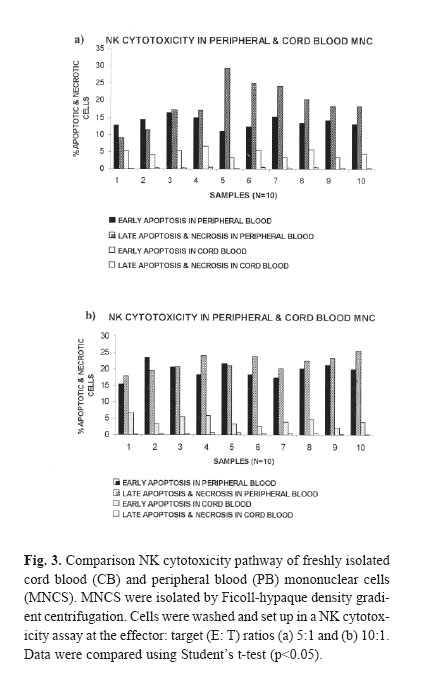
The NK cytotoxicity of cord blood (CB) and peripheral blood (PB) was determined after 4 hours of incubation in the absence of cytokines. After 4 hours in vitro incubation, co-staining with Annexin V-FITC (Axv-FITC) and propidium iodide (PI) permitted discrimination between viable, early apoptotic and necrotic cells. As we would expect, the cytotoxicity pathway in PB mononuclear cells (MNCs) consists of both apoptosis and necrosis pathways but in CB MNCs it almost consists of early apoptosis; and necrosis is negligible. With escalating E: T (effector: target) ratio changes in the percentage of apoptotic cells in PB samples were significantly higher than CB samples. The mechanism(s) of the low cytotoxicity of resting cord NK cells is not well understood. Complementary research in this field is recognized to elucidate the phenotypical and functional properties of CB cells and how they relate to maturational stages. CB studies are important for transplantation research and ma! y provide insight to the suppressive mechanism by which the host -recipient could evade GVHD and rejection. Keywords: Peripheral Blood, Cord Blood, Natural Killer Cells, Apoptosis, Necrosis, Flow Cytometry. INTRODUCTION Human natural killer (NK) cells constitute a major population of lymphocytes distinguishable from B and T cells by a lack of antigen receptor. These cells play a central role in the innate immune response in that they act both as accessory cells and as cytotoxic-effector cells, and as such, they are important in the surveillance and destruction of foreign, infected and tumor cells.1 Unlike T and B cells, NK cells do not possess a specific antigen receptor with hypervariable region and kill target cells in a non – MHC restricted manner.2 NK cytotoxicity is mediated through two mechanisms: necrosis and apoptosis. Granules within the cytoplasm of NK cells contain at least two cytotoxic mediators: Perforin, which induces necrosis through membrane damage, and granzyme B, a serine esterase that splices nucleic acids.3 Direct induction of apoptosis by NK cells was controversial until recently, though it has now gained acceptance. Although molecules involved in signaling are not yet entirely defined, CD95 may be involved.4 NK cells also secrete soluble cytotoxic cytokines such as TNFα, which induces both necrosis and apoptosis in vivo and which may be more important in the function of NK cells than either of the direct mechanisms described above.5 Cord blood NK cells produce both perforin and granzyme B. The relative role of these mechanisms in cord NK - cell cytotoxicity is uncertain.6,7 Cord blood NK cells, compared with adult peripheral blood NK cells, have a lower cytotoxic function against traditional cell lines such as K562 and Daudi.8,9 However, the low cord blood NK cell cytotoxic function can be augmented by various cytokines, such as IFN-γ, Il-2, Il-7, Il-12 and Il-15 individually and in combi-nation.8,10 Il-2, Il-12 and Il-15 raise cord blood NK cytotoxicity to the level of freshly isolated adult peripheral blood NK cells.11 However, it remains lower than the level of adult cells treated with the same cytokines.8,9 The cytotoxic activity of natural killer cells is usually tested by radioactive assay, which detects the release of cytoplasmic contents after plasma membrane disintegration of dying cells. The 51 Cr release assay is the most widely used method for measuring NK activity in vitro due to its reliability and simplicity.12 Nevertheless, as the usage of a radioactive material leads to various potential hazards, several groups have searched for easy and reliable alternatives.13 In contrast to this indirect evaluation of cytotoxicity, the assessment of cell damage by flow cytometry aims to provide a more exact characterization of the death pathway via detection of the percentage of apoptosis and dead cells.14,15 One of the characteristic processes of the apoptosis pathway is a change in the plasma membrane architecture during early apoptosis; typical membrane compounds, such as phosphatidyl serine (PS) molecules, are redirected from the inner to the outer leaflet of the cell membrane without loss of membrane integrity.15 Annexin V-FITC (Axv -FITC), a molecule with high affinity for PS, can be used to label cells in the early apoptotic state, while propidium iodide (PI) indicates late apoptosis or necrosis.16 So far, we have described a fluorescence assay that is based on the direct quantitative and qualitative flow cytometric analysis of cell damage at a single cell level. Assays were performed on 10 cord blood (CB) samples from full term normal deliveries. Results were compared with those obtained in PB samples from 10 normal adults. MATERIAL AND METHODS Effector cellsIsolation of mononuclear cells from peripheral and cord blood Cord blood samples (N=10) were obtained from full term, normal delivery, healthy infants. Human peripheral blood samples (N=10) were obtained from healthy donors (18-45 years). Samples were collected in heparinized tubes and mixed with equal amounts of phosphate - buffer saline (PBS), pH=7.4. Mononuclear cells (MNCS) were isolated by Ficoll- hypaque density- gradient centrifugation. Cell viability as measured by trypan blue exclusion was more than 99% for all cases. Cells cultured in RPMI-1640 medium supplemented with 10% fetal calf serum (FCS, GIBCO, UK), 20 mmol/L, L-glutamine, 50 U/mL penicillin, 50 μg/mL streptomycin and 20 mmol/L HEPES, referred to as complete medium .The cells were cultured at a density of 1*106 cells /mL in culture medium for the following assays. Target cellsK562 cells (erythroleukemia cell line)(Iranian Blood Transfusion Organization, Cell Bank) as target cells were grown in complete medium as stationary cultures at 37°C in 5% CO2.The cells were subcultured for 3 days before the assay, to be certain that they were in log phase. NK Cytotoxicity assayNK cell killing was assessed by modification of the method previously described.17 Target cells at the standard concentration and effector cells at various dilutions were added to create effector: target ratios (5:1, 10:1). Suspension cells were added to micro-plates and incubated for 4 hours; then, cultured cells were harvested and prepared for staining. Staining with annexin-FITC and propidium iodideAfter 4 hour co-incubation of target cells together with effector cells, total cell population was harvested from microtitre plates, collected into polystyrene tubes, then washed twice with PBS and high calcium - binding buffer (10 mM HEPES, NaOH, 150 mM NaCl, 1.8 mM CaCl2 , 1mM MgCl2 , 5mM KCl) then resuspended in 100μL binding buffer. Annexin- FITC (Axv- FITC, IQ-products, Netherlands) staining was performed with 5μL Axv- FITC and incubated for 10 min at room temperature in the dark. Immediately before flow cytometric analysis 0.1μg PI (Sigma, USA) per 100μL of the binding buffer was added and single staining using Axv-FITC or PI alone was performed as controls. FACS analysisFACS analysis was carried out on a EPICS _XL Profile cytometer (Becton Dickinson, USA) by excitation at 488nm with an Argon laser, the emission of two fluorochromes was recorded through specific band pass filter: 530nm for FITC (FL1), > 675nm for PI (FL3). Specific fluorescence was analyzed after acquiring 5000 cells. Target and effector gate and instrument variables were set according to forward and side scatter characteristics and using appropriate controls. Analysis of fluorescence was performed after back gating to exclude dead cells and aggregates. Statistical analysisAnalyses of differences within and between groups were performed using the T-test for significance (p<0.05). RESULTS Modeling for assaying apoptosis and necrosisIn order to distinguish target cells from effectors, the gating of target cells (K562) with forward and side scatter was performed. Data analysis was carried out by gating target cells followed by the analysis of Axv -FITC and PI- positive subpopulations. Axv-FITC single -positive cells (upper left quadrant) represented target cells in an early apoptotic state, Axv-FITC /PI double -positive cells (upper right quadrant) included cells in the late apoptotic and necrotic state. With escalating E: T ratios the percentage of Axv-FITC single positive or Axv-FITC double -positive cells is increased (Fig. 1, 2). For the quantification of specific cytotoxicity, the population of non -specific positive cells (determined in the control sample without effector cells) in the corresponding quadrant has to be subtracted. The proportion of corrected Axv-FITC positive events (=Axv-FITC-positive event [sample] -Axv-FITC positive events [control]) to create total events (=total events [sample]- Axv-FITC positive events [control]) × 100 reflects the real percentage of Axv- FITC positive cytotoxicity. The calculation was carried out separately for each quadrant and is exemplified for the Axv-FITC single - positive fraction. As we would expect, the cytotoxicity pathway in PB MNCS consisted of both apoptosis and necrosis pathways (in 5:1 ratios,the percentages are 13.9±1.58; 19.14±6.08 and in 10:1, 19.59±2.33; 21.86±5.63; respectively), as shown in Figure 3. However, the cytotoxicity pathway in CB MNCS almost consists of early apoptosis and necrosis is negligible (in 5:1 ratios the percentages are 4.76±2.22; 0.35±0.16; in 10:1, 5.50±1.64 and 0.32±0.16, respectively). With escalating E: T ratios in CB NK, only the percentage of cell killing by early apoptosis pathway are increased. Measurement of the early apoptosis induced by cord and peripheral natural killer cellsIn order to estimate the early apoptosis induced by natural killer cells in both cord and peripheral blood, 10 samples were evaluated. The results in Table I show that a significant variation was noticed in cord blood compared to peripheral blood at ratios of 10:1 and 5:1. As shown in Fig. 3a, the percentage of cell killing by early apoptosis pathway in PB NK was significantly higher than in CB NK. As E: T ratios escalate, the percentage of apoptotic cells in PB samples significantly increases compared with CB samples. Measurement of the late apoptosis and necrosis induced by cord and peripheral natural killer cellsIn order to evaluate the late apopotosis and necrosis in natural killer cells in both cord and peripheral blood, 10 samples were evaluated, and the results show that a significant variation was noticed in peripheral blood comparing to cord blood at ratios of 10:1 and 5:1. As shown in Fig. 3b, the percentage of cell killing by late apoptosis and necrosis pathway in PB NK was significantly higher than in CB NK. As E: T ratios escalate, the percentage of apoptotic cells in PB samples significantly increases compared with CB samples. DISCUSSIONTo date, over 1000 human umbilical CB transplants have been reported from different centers worldwide and it is generally agreed that CB represents an encouraging alternative to bone marrow (BM) transplantation.18 There are a variety of reasons for this, including the wider availability and easier access of CB as compared to BM .In addition, it has been suggested that there is a reduced graft -versus-host disease (GVHD) in CB transplants as compared to BM transplantation, even when donor and recipient are HLA-mismatched.19,22 The biological and immunological characteristics of cord blood cells differ from peripheral blood cells at several levels. Cord blood lymphoid cells are characterized by naiveté , polyclonality of T cell repertoire, and absence of certain molecules of activation, modification of cytokine production and receptors, and possible presence of suppressive cells. Whether these properties account for reduced capacity of transplanted cord blood cells to modulate a graft-versus-host disease remains to be determined. It is clear that both T cells and NK cells have reduced function when isolated from CB as compared to adult cells and both of these cell types have been implicated in GVHD pathogenesis.23,26 To support this, it has been documented that the NK cell phenotype within CB is different from those in the adult cells in that CB NK cells lack the CD57 antigen, which is found in 50-60% of adult NK cells.23 However, whether this functional and phenotypic immaturity is sufficient to explain the apparently reduced GVHD seen in CB -transplanted individuals has not yet been answered. Many studies indicate that cord blood NK activity is most likely suppressed by specific cells and others have also showed increased suppressor cells in cord blood.27 The mechanism(s) of the low cytotoxicity of resting cord NK cells is not well understood. There may be a mechanism intrinsic to cord blood NK cells, reflecting their immaturity, it may be due to extrinsic factors or it may be a combination of both. Although CB NK cells subsets were deficient in or had less lytic activity against K562 cells compared to PB NK cells, they possess the lytic molecules perforin and granzyme B and when artificially stimulated to secrete their granules during lytic assays, were capable of lytic activity equivalent to that of PB NK cells.25,26 A number of factors can modify the activity of NK cells to either increase or suppress cytotoxicity.24 Umemoto et al. showed that in contrast to fresh resting cord blood NK cells, Il-2 activated NK cells induced both apoptosis and necrosis.28 Complementary research in this field is recognized to elucidate the phenotypical and functional properties of CB cells and how they relate to maturational stages. CB studies are important for transplantation research and may provide insight in to suppressive mechanisms by which the host -recipient could evade GVHD and rejection. Here, we have described a flow cytometry-based killing assay, which permits the fine evaluation of different stages of target cell death (early apoptosis and membrane damage) occurring during NK activity responses. Target cells can be distinguished from effector cells using forward and sidescatter. Co staining with Axv-FITC and PI permits the identification of target cells at different stages of cell death [early apoptosis (Axv-FITC+/PI-) and/ or necrosis (PI+)]. These results may provide further information on the death pathway of the target cells recognized by effector cells. The assessment of cytolytic activity against target cells by Axv-FITC versus PI revealed different populations: Live cells (Axv-FITC-/PI), early apoptotic cells (Axv-FITC+/PI), late apoptotic /necrotic cells (Axv-FITC-/PI+ and /or Axv-FITC-/PI+). Since Annexin V may be bound to phosphatidyl serine located either at the outer or the inner leaflet of the plasma membrane, the direct discrimination between late apoptotic and necrotic cells is critical. With increasing E/T ratios, we observed an accumulation of two different populations of target cells (Fig. 1,2), the majority consisted of Axv-FITC+/PI- early apoptotic cells while the minority were found to be Axv-FITC-/PI+ late apoptotic target cells. No significant increase in Axv-FITC-/PI+ necrotic cells was observed after 4 hours of incubation. These data suggest that most of the target cells recognized by effector cells die via the apoptosis pathway. The flow cytometry assay has advantage over the 51 Cr release assay; the assay may measure either apoptotic or necrotic cell death of the target cells. Perforins are known to create channels in the cell membrane through which granzymes released by NK cells can induce the apoptotic cascade.28 Therefore, some targets die a necrotic death from perforin lysis, while other cells die a slow, apoptotic death. The 51Cr release assay measures the 51Cr released by necrotic cells, whereas apoptotic cells may not release 51Cr. In conclusion, the flow cytometry assay is a quantitative method for the detection of cell- mediated cytotoxicity and does not use radioactive labeling .The reliability and reproducibility of this assay make it a useful alternative to the standard 51Cr release assay, providing a greater insight in to the mechanisms of cell-mediated cytolysis by different effector populations. REFERENCES
Copyright 2004 -Medical Journal of the Islamic Republic of Iran The following images related to this document are available:Photo images[mr04009f1.jpg] [mr04009f2.jpg] [mr04009f3.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}