 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
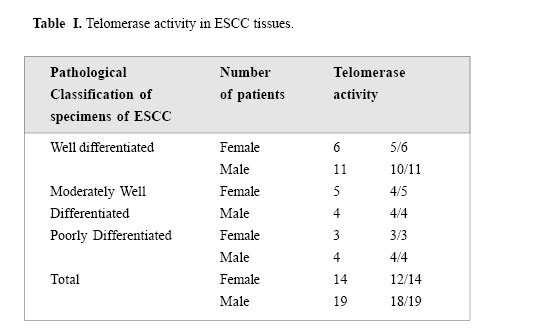
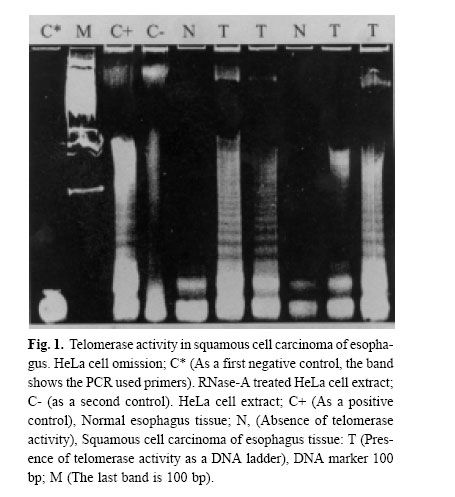
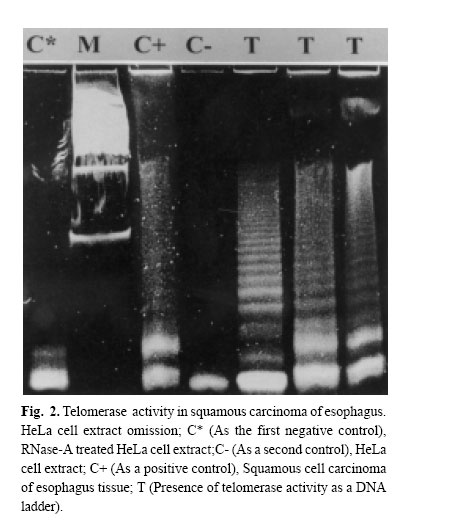
Medical Journal of the Islamic Republic of Iran , Vol. 18, No. 2, August, 2004, pp. 147-152 TELOMERASE ACTIVITY IN IRANIAN PATIENTS WITH ESOPHAGEAL SQUAMOUS CELL CARCINOMA S. KAZEMI NOUREINI,* A.A. ZIAEE,* Ph.D., F. RASTEGAR-JAZII,+ Ph.D., AND M. YAZDANBOD, M.D.++ From the *Institute of Biochemistry and Biophysics, P.O.Box: 13145-1384, University of Tehran, Tehran, the +National Research Center for Genetic Engineering and Biotechnology (NRCGEB), Tehran, and the ++Department of Surgery, Shariati Hospital, Tehran University of Medical Sciences, Tehran, Iran. Correspondence: AA. Ziaee, Institute of Biochemistry and Biophysics, P.O. Box 13145-1384, University of Tehran, Tehran, Iran; E-mail: aa_ziaee@yahoo.co.uk. Code Number: mr04028 ABSTRACTTelomerase activation is one of the main pathways to immortalize cancer cells. In many kinds of cancer cells, this special reverse transcriptase stabilizes and elongates telomeres and prevents telomere erosion that naturally occurs in every cell division. Esophageal cancer is the fifth most frequent cause of cancer death worldwide, and is highly associated with alcohol, smoking, cultural habits, and environmental factors. Telomerase has been suggested as a tumor marker and a molecular target for drug design in several kinds of cancers. In this work telomerase activation was inspected among Iranian patients with Esophageal Squamous Cell Carcinoma (ESCC), and detected in 90% of samples of different stages. This may be an indication that telomerase activation happens in an initial step in the development of ESCC. Although there is no correlation between telomerase activity and the progress of ESCC, it could be considered as a good tumor marker in ESCC. Telomerase activity te! sts are suggested for screening purposes in high risk areas for ESCC, which can be easily done on a small amount of scrapped samples of esophageal mucosa. It is also possible that ESCC results from incomplete differentiation or a failure in telomerase gene switching off that normally occurs during the differentiation of esophageal epithelial cells. Keywords: telomerase, telomere, esophageal squamous cell carcinoma, TRAP assay. INTRODUCTIONEsophageal cancer is ranked as the fifth cause of death by cancer in the world.1 The prognosis for both types of esophageal cancer, squamous cell carcinoma (ESCC) and adenocarcinoma (ADC), is poor; a survival rate at 5 years of less than 10% has been reported.2 In developing countries, more than 85% of cases of esophageal cancer are ESSC, associated with a variety of significant factors coming from their environment, cultural and nutritional habits.3,4 Different pathways may be involved in the pathogenesis of esophageal cancer. Actually, one of the most intriguing aspects of the epidemiology of esophageal cancer is its strong geographical variation; certain areas of the world like the north parts of China, around the Caspian sea, South Africa and South America have a higher incidence by several orders.5 Although the mortality from esophageal cancer has decreased in the world using new and improved diagnostic strategies by endoscopy and microsurgeries, the 5-year survival rate of patients with resectable esophageal cancer after treatment ranges between 20 and 30%, mainly because the majority of patients with esophageal cancer are diagnosed in late stages. Recent molecular oncology studies on Iranian patients with ESCC performed in our lab have shown that 61% of Iranian patients with ESCC had a loss of heterozygosity (LOH) of a region on 17p, around the location of p53 gene (telomeric region), whereas 59.4% showed an allelic loss of 17q (Ziaee et al., unpublished data). It is noteworthy that the telomere length of chromosome 17p is shorter than median telomere length, suggesting chromosome-specific factors regulate telomere length. Therefore, it is possible that the short telomere of 17p and frequent loss of alleles in ESCC tumors6 may influence tumor cell behavior. Telomerase as a ribonucleoprotein, consists of an internal RNA, catalytic protein component and several other auxiliary factors to extend the 3'-G-rich strand of the ends of the telomeres. This enzyme is largely absent in normal somatic cells, while it is activated in immortal and cancer cells.7 Telomere bound hnRNPs may prove to be good targets for regulating telomere length.8 Human telomerase (hT) adds repeated units of the 5'-TTAGGG-3' hexamer to the ends of telomeres by reverse transcription activity, using its RNA component (hTR) as a template9 and compensates the telomere loss that normally occurs in every cell cycle by the incomplete replication of linear chromosomal DNA.10 Telomere crisis, which is a result of continued unregulated cell division, promotes secondary genetic alterations that enhance tumorigenesis and tumor growth is facilitated by telomerase activation (for a review see).11 On the other hand, very short telomeres are protected and stablized by an unknown mechanism. So telomerase acts more than just elongating the telomeres, it is also under regulation in multiple levels; regulation of hTERT transcription, transport to the nucleus, assembly of the telomerase holoenzyme, recruitment of telomerase to the telomere, and post-translational modifications of hTERT protein, although generation of full-length hTERT protein is probably the limiting step.12 In many different cancer cells, telomerase is activated and it is considered as a very good tumor marker. Telomerase activation is achieved at various steps, including transcriptional and post-transcriptional levels of the hTERT gene.13 The expression of hTERT in tumor cell lines shows a recessive phenomenon, indicating that lack of expression in normal cells is because of one or several repressors.14 Several chromosomes may code for such repressors including some transcription factors, cell cycle inhibitors, tumor suppressor genes, hormone receptors, cell fate determining molecules and viral proteins.14 However, no clear explanation has been outlined so far in connection with the tumor specific expression of the hTERT gene.14 Telomerase activity have been detected in a considerable number of esophageal carcinoma and also normal esophageal mucosa.15 In 1998, Asai et al. studied 13 human ESSC cell lines, and showed telomere restriction fragment (TRF) shortening and telomerase activity in all of these cell lines.16 Applying in situ hybridization technique, hTR overexpression has been reported as an early event in the development of ESCC.17 From the molecular oncology point of view, studies on telomerase activity in Iranian patients with ESCC can be a valuable contribution to the efforts of elucidation the esophageal cancer etiology in high risk areas around the world. MATERIAL AND METHODSFresh tissue samples of ESCC were snap frozen and kept in liquid nitrogen until use. Histopathological studies of tissues were performed according to TNM classification by two pathologists blind to the telomerase activity results of the tissues. Tissue samples were powdered by microdismembrator at maximum speed for 60 seconds under liquid nitrogen conditions. Each powdered tissue sample was added to an appropriate amount of lysis buffer containing 10 mM Tris-HCl pH=7.5, 1 mM MgCl2, 1 mM EGTA, 0.1 mM Phenylmethylsulfonylfluoride (PMSF), 5 mM betamercapto ethanol, 0.5% CHAPS and 10% glycerol.25 After 30 minutes incubation on ice, the lysate was centrifuged at 16000g for 30 minutes at 4°C. Supernatant was then applied for Telomerase Repeat Amplification Protocol (ATRAP) assay.25 Due to the well known presence of the telomerase activity in HeLa cells, these cells were used as a positive control. If HeLa cellular extract is treated with RNase-A ribonuclease, it can be consid! ered as a negative control as well. HeLa cells were purchased from Iran Pasteur Institute (ATCC No: CCL-2). Initially, HeLa cells were grown in in RPMI 1640 medium supplemented with 10% fetal calf serum (FCS) and 100 units Penicillin and incubated at 37oC under 5% ambient COcondition. For cellular extract preparation, cells were grown to 80% confluency, trypsinized with 0.2% trypsin, washed with phosphate buffer saline, centrifuged then resuspended in lysis buffer at a concentration of 5*106 cells/mL. Protein concentration of all samples were estimated using a Bradford based microassay.38 For the telomerase assay, 0.05–5µg of the extracted protein were used for a 50 µL reaction mixture containing 20 mM Tris-HCl pH=8.3, 1.5 mM MgC12, 63 mM KC1, ImM EGTA, 0.005% Tween 20, 0.1 mg/mL BSA, lug T4g32pr, 20 µM dNTP mix), 0.1µg Ts primer (5'-AATCCGTCGAGCAGATT-3') and after incubation at 24-26°C for 20 minutes 0.1 µg Cx primer (5' -(CCCTTA)4 -3'), the product of reverse tra! nscriptase activity of the telomerase was ready for PCR amplification. The PCR amplification consisted of an initial denaturation, without Taq polymerase, at 94 °C for 5 min, then 2 units of Taq polymerase were added and 30 cycles at 94°C for 30 s, 52°C for 30 s and 72°C for 45s were performed on a Techne Thermal Cycler. Controls for telomerase activity TRAP assay, consisted of both 0.5 µg RNase-A ( for destructing the RNA component of the telomerase) treated and untreated HeLa cellular extract as negative and positive controls, respectively. Another control containing the PCR mixture without any cell or tissue extract was used to check primer dimerisation and any sort of genomic contamination. The PCR product was electrophoresed on 15% non-denatur-ing polyacrylamide mini-gel, stained with silver nitrate. RESULTSIn this study 33 ESCC cases were examined. A tumorous part and an adjacent normal tissue as a normal pair were taken from each ESCC patient. Tumors were then classified in 3 major groups by a pathologist considering TNM classification: well differentiated, moderately well differentiated and poorly differentiated ESCC. Looking at Table I, one can see that the average age to develop a tumor is about eight years lower for women than for men. However, the quantity of samples analyzed is too low to statistically reflect the whole population of ESCC patients in Iran. TRAP assay showed telomerase activity at least in 30 out of 33 samples (90%). This high frequency of telomerase positive tumor tissues suggests that activation of this enzyme happens in an early stage in tumorigenesis of ESCC and stay switched on during tumor progression (Figs. 1,2). Our data analysis indicated no correlation between the telomerase activity of samples and the stage (or grade) of esophageal squamous cell carcinoma, the prognosis, sex, age and geographical distribution of patients who were from different long distant areas in the country. There was a very slight telomerase activity observed in some normal adjacent epithelial tissues of esophagus, which is not comparable with the telomerase activity of their tumor pairs. This small amount of telomerase activity in normal tissues is well understood to be due to the presence of some stem cells which naturally proliferate and regenerate the eplithelium of normal esophagus and/or some small contamination with tu! mor cells in the apparently normal parts of esophagus adjacent to the tumor, which has been considered as normal tissue when inspected by bare eyes during tissue processing. DISCUSSIONAlthough telomerase activity can be easily studied using RT-PCR of RNA component of telomerase (hTR),18 and reverse transcriptase subunit of telomerase (hTERT),19 it has frequently been preferred to determine telomerase activation on the basis of catalytic activity of this enzyme. To evaluate such activity, TRAP, PCR-ELISA or other common assays were used as these are more reliable and perfect and recommended experiments. However, several publications have shown the complexity of the telomerase,20,12 which implicate the complicated regulation of telomerase activity. For example, at the gene expression level, different isoforms of this enzyme result following alternative splicing of the same mRNA, and some of which are catalytically inac-tive.21 This in turn can titrate the hTR component in the RNP complex of telomerase and lead to decrease in activity. Post-transcriptional modification has also been reported and inhibitor pathways may exist. The introduction of a normal human chromosome 3 into human renal cell carcinoma caused repression of telomerase activity,22 and protein phosphatase 2A can inactivate telomerase.23 hTR component has also been seen without any detectable telomerase activity in certain human normal and tumorous cells and several preimmortal cells. In some transgenic mouse models, the RNA component of telomerase was up-regulated in early preneoplastic stages, while the telomerase activity is only lately ob-served.24 In our study, we preferred to evaluate telomerase activity by the well recommended TRAP assay, with slight modifications of Kim et al’s protocol.25 We used fresh tumors without considering the degree of ESCC development, and telomerase activity was detected in about 90% of the tumor samples. The molecular changes during esophageal carcinogenesis remains to be understood, however many genes are deregulated in ESCC.26 COX-2 overexpression and the mutations profile of p53 gene in ESCC Iranian patients recently has shown a high prevalence of C to T transition at CpG dinucleotides in about 50% of tumors suggesting a baseline of chronic inflammatory stress involved in ESCC in this high risk area of incidence.27 The higher incidence of telomerase activity in this work to other events such as p53 mutations and/or LOH suggest ! that the occurrence of such telomerase activity may be a critical step in ESCC carcinogenesis. Considering recent data on telomere structure, telomere-binding proteins, and sensitivity of telomeres to oxidative damage, a new model suggests that t-loop formation prevents access of repair proteins to telomeres, and this in turn may lead to the accumulation of apurinic and apyrimidinic sites and single-strand breaks that accelerated telomere shortening. In consequence of this, the single-stranded G-rich overhang, which is normally protected at the base of the telomeric loop, would be exposed to the nucleoplasm and act as a strong inductor of the p53 pathway. Such exposure of the overhangs has been proposed to be the first step in the signal transduction cascade to replicative senescence.28 Cells lacking normal activity of p53 may pass senescence easier and become more potent in proliferation. Epidemiological studies have indicated that environmental factors such as working place pollution including chemicals, corrosive, fine particles,29 and some cultural habits in food preparation, and kinds of foods of interest in high risk areas of incidence are associated to esophageal cancer. These factors may play an important role causing frequent microinjuries in individuals with prolonged exposure. Therefore it is possible to suggest that ESCC results from incomplete differentiation of esophageal epithelial cells during the development of these microinjuries or a failure in telomerase gene switching off normally occurs during the normal differentiation of esophageal epithelial cells. ESCC shows a high rate of incidence in developing countries and usually it is not diagnosed until the growing tumor prevents mouthful swallow, which is inevitably ended by at least partial resection of this part of the digestive tract in patients who also develop anorexia. Proposing telomerase activity as a good tumor marker for this kind of cancer would be a useful tool for acceler ating its diagnosis. This assay may easily be performed in combination with endoscopic inspection or together with screening studies without resecting biopsies and probably just by esophageal washing and/or a mild scrape of the epithelial tissues of the esophagus, This may be similar to the recently reported telomerase activity in cytologic fluids.30 On the other hand telomerase may be considered as a good target for chemotherapy31 and immunotherapy32 in cancer treatment. However, the presence of telomerase activity in stem cells or germ cells, which seems necessary for their progeny, may causes uneasiness with the use of telomerase inhibitors. Considering that the telomeres length in normal proliferative cells are much longer than tumor cells and most primitive stem cells that occasionally proliferate, their telomeres should be shortened at a much slower rate.33, 34, 35 These physiological aspects have led many investigators to focus on designing telomerase inhibitors for anti-cancer chemotherapy protocols.31 It has recently been reported that inhibition of telomerase activity by peptide nucleic acid and 2’-O-MeRNA oligomeres, caused reversible progressive telomerase shortening and induced apoptosis in all those treated immortal human cells that finally led to their death. When the supplement of these inhibitors we're stopped, telomeres regain their initial lengths.36 Also some other chemicals such as oligonucleotide N3'→P5' phosphoramidates, and thio-phosphoramidates have recently been checked for having such inhibitory prop-erties.37 Such updated achievement gives hope for a therapy allowing non-cancerous cells to regain their health after chemotherapy. ACKNOWLEDGEMENT The authors acknowledge the Research Council of University of Tehran for the financial support under project No. 521/5/662. We are grateful to Jacques Poudrier for the critical reading of this manuscript. REFERENCES
Copyright 2004 -Medical Journal of the Islamic Republic of Iran The following images related to this document are available:Photo images[mr04028f3.jpg] [mr04028f2.jpg] [mr04028f1.jpg] [mr04028t1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}