 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
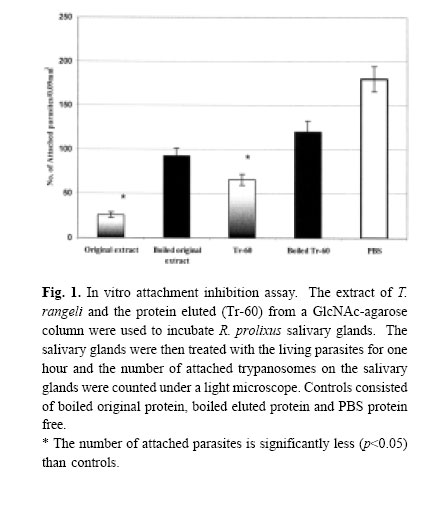
Medical Journal of the Islamic Republic of Iran , Vol. 18, No. 2, August, 2004, pp. 159-166 IN VITRO INHIBITION OFATTACHMENT OF THE AMERICAN TRYPANOSOME, Trypanosoma rangeli ON THE SALIVARY GLAND OF ITS VECTOR, Rhodnius prolixus, USING THE PARASITE EXTRACT AND PARTIAL PURIFICATION OFAN ATTACHMENT INHIBITOR PROTEIN H.R. BASSERI, Ph.D.,* AND N.A. RATCLIFFE ** From the *Department of Medical Entomology, School of Public Health, Tehran University of Medical Sciences, P.O. Box 14155-6446. Tehran, Iran, and the **Biomedical and Physiological Research Group, School of Biological Sciences, University of Wales Swansea, Singleton Park, Swansea SA2 8PP, United Kingdom. Correspondence: H.R. Basseri, Department of Medical Entomology, School of Public Health, Tehran University of Medical Sciences, PO Box 14155-6446. Tehran, Iran. E.mail: HYPERLINK“mailto:Hamid_basseri@hotmail.com” Hamid_basseri@hotmail.com Code Number: mr04030 ABSTRACTThe life cycle of the American trypanosome, Trypanosoma rangeli, in its invertebrate host, Rhodnius prolixus, is completed by invasion into the insect salivary glands. As a prerequisite, there is an initial recognition of the salivary gland receptors for the adhesion/invasion by T. rangeli. Therefore, the purpose of this study was to investigate the putative receptor molecules on either the salivary gland or parasites surfaces, which mediate the adhesion/invasion. Initially, the possible agglutination activity of T. rangeli epimastigotes against blood cell was examined. Then, in order to investigate the role of any receptor ligands on the surface of T. rangeli in adhesion/invasion of the parasite, GlcNAc was preferentially used to isolate a ligand protein. A 60kDa protein bound to the GlcNAc column and was isolated from T. rangeli (Tr-60) and the purity of this protein was confirmed by reverse phase HPLC. Tr-60 also exhibited a potent parasite inhibitory effect indicating! that this protein (most probably a glycoprotein) may act as a ligand receptor. In vivo, the development and invasion of T. rangeli into the salivary gland was not interfered with by the presence of a sugar inhibitor of the R. prolixus hemolymph lectin. Keywords: Trypanosoma rangeli, Rhodnius prolixus, Salivary glands, Proteins, attachment. INTRODUCTION T. rangeli needs an essential period of development before invasion into the salivary gland and the long epimastigote forms are responsible for invasion.5 The adhesion inhibition of T. rangeli onto R. prolixus salivary glands using sugars revealed that some carbohydrate- binding proteins on the surface of the trypanosome or gland acted as ligands mediating the parasite adhesion.5 Therefore, in this study, a sugar binding protein was used to purify some ligand proteins. Parasite adhesion/invasion is likely to be prefaced by specific receptor-ligand interactions between moieties on the cell surface of the parasites and molecules of vector tissues.19 Such surface receptors are beginning to be characterized for Trypanosoma cruzi. In this parasite, three life-cycle stages are characterized by isolating glycosylated proteins from their cell-surface24 and inhibition of T. cruzi invasion in cell culture by antibodies strongly suggested that at least one of these polypeptides could be the receptor for T. cruzi in the host cell1 . A glycoprotein on the surface of T. cruzi is involved in adhesion/invasion of the T. cruzi to the mammalian cell body.19 This glycoprotein with molecular weight 82kDa, is expressed in the trypomastigotes of T. cruzi15 and it is also essential for physical movement and natural transmission of T cruzi. 8 In addition, the evidence for the invasion of the African trypanosome, Trypanosoma brucei, within the invertebrate host would suggest that they have some means of recognising and adhering to one or more ligands in the vector.14 Procyclic molecules (glycoproteins on the surface of the epimastigote forms of the parasite) allow the trypanosome to bind to lectin type molecules within the vector.21 Lectins play a role in attachment of some parasites to the host cells, e.g. a lectin was found in Entamoeba histolytica which mediates in adhesion of trophozoites to monolayers of human cells.6 The other possible function of lectins in protozoans include binding to red blood cells by Plasmodium falciparum,10 cell-to-cell interaction with Cryptosporidium parvum29 and determination of the particular site of infection with Eimeria species.25 Chatterjee et al., (2003) found a lectin-like component with broader specificity on the surface of L. amazonensis promastigotes. Schottelius (1992) also reported the presence of lectin activity in promastigotes of 18 strains of Leishmania but not in amastigotes and, subsequently, Svobodova et al., (1997) detected hemagglutination activity in 16 strains of eight Leishmania species. In addition, they reported that the incubation of lysates in temperatures up to 100°C had no effect on hemagglutination activities. This study was conducted mainly in UK and partially Iran in 2001-2002 and the purpose of it was to investigate the role of receptor ligands on the surface of T. rangeli in attachment/invasion of the parasite. In particular, the inhibitory effect of the parasite’s protein extract to interfering with specific steps in the invasion of the salivary glands has been examined. In previous study it has been shown that GlcNAc showed high inhibitory effect particularly to the sugar pre-incubated parasite.5 Thus, GlcNAc-binding protein as an example may help to further clarify the mechanism of parasites’ invasion of the host cell. From this point of view, in the present study, it has also been tried to purify and characterize this receptor from the surface of T. rangeli long epimastigote form. MATERIAL AND METHODS Parasites cultureT. rangeliTejera 1920, strain Choachi was supplied by Dr. G. Schaub (University of Bochum, Germany). Trypanosomes were maintained at 25° C in 10mL sterile centrifuge tubes containing LIT culture medium [0.068M NaCl, 0.0054M KCl, 0.056M Na2 HPO4 , 0.01M glucose, 5mg ml-1 tryptose, 5mg mL-1 LIB (liver infusion broth, DIFCO), and 0.0025mg ml-1 haemin (Sigma Co., Dorset, UK)], supplemented with 20% heat-inactivated fetal calf serum (FCS) (Takle and Young 1988). Insect and dissection of salivary glandsR. prolixus was reared as described by Azambuja and Garcia (1997) and fed through a parafilm membrane on defibrinated horse blood (TCS Microbiologicals Ltd, Botolph, Claydon, Buckingham, UK) every 10-15 days and occasionally with rabbit blood (Harlan Sera Lab Ltd, Loughbrough, UK) or fresh human blood. The salivary glands, the insects were anesthetized on ice for 10 min and then their heads were held firmly with No. 5 forceps (BIS Switzerland) and immersed in 10mM PBS (containing 150mM NaCl and 10mM sodium phosphate at pH 7.4 and 380 mOsm) or TBS (100mM Tris, pH 7.3, 300 mOsm, 150mM NaCl). The head and abdomen were then gently pulled apart. This allowed access to the salivary gland ducts, which remained attached to the head. The glands were then separated from the head. Preparation of T. rangeli extractsThe epimastigotes long forms of T. rangeli from three week and short forms from one week old LIT medium were harvested and washed twice with PBS (containing 150mM NaCl and 10mM sodium phosphate at pH 7.4) by centrifugation at 10000g IEC (Centra-M centrifuge, International Equipment Co. USA) for 10 minutes. The pellet was resuspended in 100µL PBS to give a final concentration of 2.5-4.5 *107 cells/mL. Because of hard membrane and base on previous tests, the suspension of parasites was homogenised with an ultrasonic disintegration sonicator (Ultrasonic Disintegrator150 watt, MSE, UK) for 1 minute and then spun down at 10000g for 10 minutes and the extracts were frozen at -20° until use. Hemagglutination assayTwo-fold serial dilutions of long and short epimastigote form extract were prepared in sterile 60 well microplates (5 µL) (Nunc, Gibco, U.K.) to give a final dilution range of 1:2 to 1:1024. Five microliters of 2% rabbit erythrocytes in PBS were added to each well and the mixtures were agitated for 1 minute and then incubated for 2 hours at room temperature. The end points of agglutinin were examined under a stereomicroscope and by the flow characteristics of the erythrocyte pellets when the plate was held at an angle. As controls, 100µL of the extract was heat-inactivated for 20 minutes at 100° C. All samples were prepared 3 times and the assay repeated each time. Attachment inhibition assay using T. rangeli extractThe salivary glands of 4th and 5th instar nymphs of R. prolixus (4 glands each time) were dissected as above and then preincubated with the extract of T. rangeli for 1 hour at room temperature. The salivary glands were then quickly washed and incubated with the living trypanosomes (2*106 cells/mL) in PBS for 6 hours at 25° C. Each salivary gland was washed in PBS and the attached trypanosomes were counted by counting from the upper surface area of the posterior and anterior lobes of the salivary glands perpendicularly to the viewing direction, using 100 squares in an eyepiece grid graticule measuring 0.09 mm2 . Controls consisted of the same number of the salivary glands that had been preincubated with heat-inactivated trypanosome extract and untreated salivary glands in PBS. The experiment was repeated five times with 2 replications each. Partial purification of a T. rangeli attachment proteinT. rangeli epimastigote long forms were harvested from a large volume (5000 mL) of 3 week old LIT culture medium by centrifugation at 14000g (High Speed 18, MSE, UK) for 30 minutes at 4° C. The pellets were resuspended in 10mM PBS (pH 7.4; 380mOsm), and washed three times by centrifugation at 10000g for 15 minutes each and then resuspension in 5 mL PBS containing 1% sodium dodecylsulphate (SDS). Trypanosomes were homogenized with an ultrasonicator (Ultrasonic Disintegrator150 watt, MSE, UK) and centrifuged at 10000g. The supernatant was then applied to a GlcNAc-agarose column for protein purification. Five millilitres of the supernatant were loaded onto a 1mL GlcNAc-agarose column (Sigma Co. Dorset, UK) by adding 1mL supernatant each time and incubating for 1 h each at RT. The column was then washed with 20 mL PBS pH 7.4, followed by 20 ml of the buffers containing 0.5M and 1M NaCl each, and finally with 20mL of PBS pH 7.4. The bound proteins were eluted using 5mL of 1 mg/mL GlcNAc. The eluted protein was concentrated and washed with the buffer to remove the GlcNAc using a Centricon 30 concentrator (Amicon Inc. Beverly, USA) at 1000 g at room temperature. The sample was in vitro attachment inhibition assays as above, SDS-polyacrylamide Gel Electrophoresis and Reverse- phase High Performance Liquid Chromatography (HPLC). SDS-Polyacrylamide Gel ElectrophoresisThe eluted protein was applied to SDS-PAGE as discussed by Laemmli (1970). Twenty microliters of T. rangeli attachment protein was concentrated 5 times by freeze-drying (approximately 0.2-0.4 mg/mL), and, subsequently, in order to denature the protein, the protein was mixed with 5µL of 5x SDS-PAGE sample buffer under non-reducing conditions and then heated for 10 minutes at 100° C. The sample (20µL) was then centrifuged at 10000g for 5 min to remove debris before loading into the gel. The supernatant was applied to SDS-PAGE electrophoresis in a 10% polyacrylamide gel to analyse the eluted protein. Electrophoresis was performed at a constant voltage of 200v for ca., 45 min using the bio-Rad mini-protean II apparatus (Bio-Rad laboratories LTD., Hemel Hempstead, Hertforshire, UK). After electrophoresis, the gels were fixed with 40% ethanol plus 10% acetic acid for 40 minutes. Detection of the protein band was improved by staining the gel with 0.2% (w/v) silver nitrate as de! scribed by Marshall (1984). Finally, the gel was developed in 2.5% (w/v) sodium carbonate containing 0.04% (v/v) formalin. For calibration of molecular mass, the wide range precision protein standard (pre-stained) marker kit (Bio-Rad) was used and the actual molecular weight was measured by GenGenius Gel Documentation and Analysis System (Syngene, Cambridge, UK). High Performance Liquid Chromatography (HPLC)HPLC analysis was carried out using a LDC/Milton Roy apparatus consisting of a spectromonitor D, constametric pumps and a CI-4100 or CI-10P integrator, using a Spherisob-ODS C18 analytical column (25*0.34cm) at a flow rate of 1 mL/min to check purity of the eluted protein from the GlcNAc-agarose column. The mobile phase was 50% CH3 CN: 50% H2O containing 1% trifluoroacetic acid (TFA) (Sigma Co., Dorset, UK). Ten microliters of the eluted protein was injected and the purity of the protein solution was detected at a wavelength of 220 nm. RESULTS Hemagglutination assayThe trypanosome extract showed a weak hemagglutinin activity (titre< 16) against rabbit red blood cells. In contrast, no hemagglutinin activity was observed in the extract of short epimastigotes and of the eluted protein. The heat treatment of the long epimastigote extract at 100°C did not completely destroy the agglutinin activity (Table I). Attachment inhibition assay using T. rangeli extract.The extract of T. rangeli epimastigote long forms showed a high inhibitory effect (85.7% reduction in comparison with the PBS control) on the attachment of live parasites to the surface of R. prolixus salivary glands
*The control was inactivated by heating at 100oC for 20-30 minutes. ND, not done. Table II. In vitro attachment inhibition assay. An extract of T. rangeli was loaded onto a 1ml GlcNAc-
Agarose column. GlcNAc-binding protein (Tr-60) was eluted using 1mg/mL GlcNAc in 10mM PBS.
The eluted protein (Tr-60) as well as original extract were used to treat the Rhodnius Salivary glands and
washed. The salivary glands were then treated with the living parasites for one hour and the number of
attached trypanosomes on each salivary gland was counted under a light microscope.
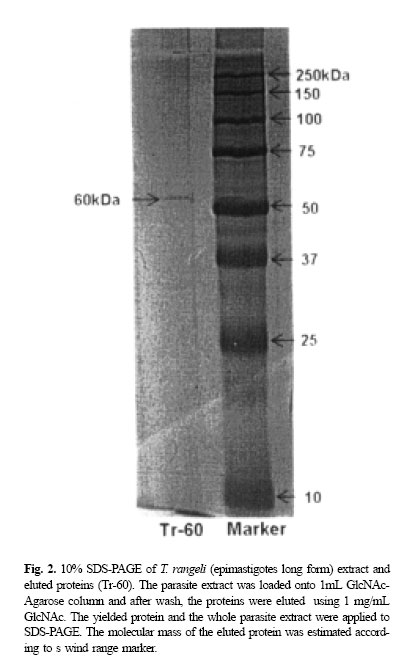
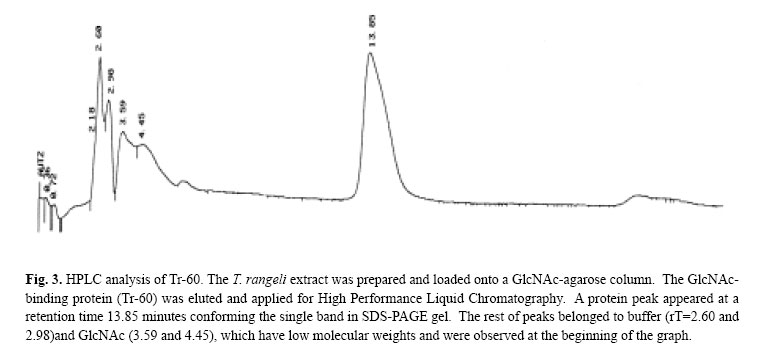
while the protein eluted, (Tr-60), showed 63.5% reduction in parasites adhesion which was significantly different from the original protein compared with the PBS control. The inhibitory effect was not completely destroyed when both original extract and Tr-60 were heated at 100oC (Table II). The heat-treated original protein reduced adhesion by 49.2% of the trypanosomes while the reduction for the heat-treated Tr-60 was 33.6%. However, the number of trypanosomes attached to the salivary glands pretreated with the original extract was significantly (p< 0.05) different (71.6% reduction) compared with its boiled control (Fig. 1). Tr-60 also showed a significant reduction (45.2%) in parasite adhesion in comparison with heat-treated Tr-60 (Fig. 1). Above all, attachment in the PBS protein free preparations was significantly higher (p< 0.05) compared with the trypanosome extract, eluted protein and their boiled controls. SDS-Polyacrylamide Gel Electrophoresis and HPLCThe molecular mass of the eluted protein was estimated according to the wide range marker ca. 60kDa (Tr60) (Fig. 2) while the whole protein extract loaded on the gel, showed numerous bands. The purity of the eluted protein (Tr-60) was also tested by reverse phase HPLC (Fig. 3). When this protein solution was injected into the HPLC analytical system, a peak appeared at the retention time of 13-14 minutes (rT=13.85) after loading (Fig. 3) which was 51.6% of the area of the graph. The remaining peaks belonged to PBS (rT= 2.60 and 2.98) and GlcNAc (rT=3.59 and 4.45) which have low molecular weights and were observed at the beginning of the graph. DISCUSSIONThe in vitro adhesion inhibition assay using extracts of long epimastigotes of T. rangeli revealed significant adhesion inhibition by blocking the receptors on the salivary gland surface. This process was specific to the extract from the epimastigote long form as no such inhibition occurred with the short form of the parasites. These results indicate that T. rangeli long epimastigotes possess the appropriate ligands for binding to R. prolixus salivary glands, which mediate the initial attachment of the parasites. In fact, in each parasite stage, specific molecules associated with the surface membrane are most probably involved in interactions with the hosts.8 As previously has been described,5 in the invertebrate phase of T. rangeli, only long epimastigotes can attach to the surface of R. prolixus salivary gland and, this process can be blocked by pre-incubation of parasites or salivary glands with specific sugars.5 Several functions have been ascribed to sugar-bind-ing proteins on the surface of protozoans,11 one of the main roles being the interaction between these molecules as receptors with host cell surface carbohydrate, during parasite attachment/invasion.20 One of the candidates for the attachment molecules may be lectins which are known to mediate cell-to-cell interaction.23 These molecules have been reported from several parasitic protozoans such as Leishmania species promastigotes and amastigotes,26 Plasmodium falciparum,13 Tritrichomonas foetus 3 and Entamoeba histolytica . 28 In the present study, the extract of long epimastigotes of T. rangeli showed low agglutinin activity against rabbit blood cells so that the attachment molecules may well be lectin-like in this parasite. The effect of carbohydrate inhibitors on attachment of some protozoans to their host cells suggests the presence of lectin or lectin-like molecular activity in the Protozoa.20 Several authors have studied surface protein receptor in protozoan adhesion. For example, the 82kDa glycoprotein on the surface trypomastigotes of T. cruzi which is involved in mammalian cell invasion.23 Jungery (1985) also reported that P. falciparum merozoites had lectin-like proteins on their surface that bound to red blood cell carbohydrate receptors and the merozoite invasion in vitro could be specifically blocked by GlcNAc, GalNAc and NeuNAc. Recently, three merozoite surface proteins have been identified that are specific for GlcNAc binding10 and could be involved in the binding of merozoites to the glycoconjugates on the surface of the red blood cells. It has been shown the presence of six sugars on the surface of R. prolixus salivary glands5 and also those sugars were employed for the attachment inhibition assays in vitro by authors. The results showed T. rangeli adhesion was blocked strongly by GlcNAc indicating the presence of specific ligand on the parasites and salivary gland. Therefore, on the hypothetical possibility that the attachment protein may bind to a sugar affinity column, in the present study, GlcNAc was preferentially chosen to purify a ligand protein from T. rangeli using a GlcNAc-agarose affinity column. The isolated protein has apparent molecular mass at 60 kDa by SDS/PAGE and its purity was confirmed with HPLC (Fig. 3). This protein even at low concentration (0.083 mg/mL), showed attachment inhibitory effect (Fig. 1) suggesting that T. rangeli receptors specifically interact with glycosylated salivary gland surface molecules such as GlcNAc moieties which are present on the salivary gland. In additio! n, the results showed that the molecule causing the inhibitory effect from both the trypanosomes extract and the Tr-60 were not completely heat-sensitive. WGA-labelled lectin has a high affinity for GlcNAc, exhibited strong and specific labelling of the salivary gland basal membrane and cell layer.5 This interaction may confirm the previous finding, indicating the high inhibitory effect of pre-incubating parasites in GlcNAc and subsequent experiments, described here, show the interaction of this sugar with protein receptors on the surface of T. rangeli. This interaction was not affected by extraction of parasites using solvent, indicating that the binding to Tr-60 was mediated by glycoproteins rather than glycolipids. Generally, these results strongly show the presence of carbohydrates on the cell-surface of R. prolixus salivary gland, which are linked to glycoproteins by N-glycosylation. Several authors have reported the role of glycosylated proteins in invasion of parasitic protozoans into the host cells. For example, Barreau et al., (1995) reported the presence of carbohydrate-binding proteins on the surface of the salivary gland of Aedes aegypti serving as receptors for invasion of the sporozoite of Plasmodium gallinaceum. The role of carbohydrate-mediated adherence has also been shown in a mannose-binding protein of Acanthamoeba12 and a galactose/GalNAc-binding protein of Entamoeba.9 Finally, in the present study, the 60 KDa protein isolated from T. rangeli epimastigote long form can recognize GlcNAc specific receptor present on R. prolixus salivary glands and are involved in the parasites adhesion. In addition, an attempt to find the sequence of Tr-60 using Edman chemistry failed probably due to modification of the N-terminus of the protein. CONCLUSIONExperiments described in this study, show the blocking of the attachment of T. rangeli long epimastigotes in vitro to the surface of R. prolixus salivary glands using the extract of trypanosomes. Also, the presence of low agglutinin activity in the parasites extract may indicate the epimastigote long form possesses lectin-like molecules. GlcNAc, which showed the highest inhibitory potency with respect to blocking the trypanosome adhesion on the salivary gland surface, was preferentially used to purify a ligand protein of T. rangeli. A 60 kDa protein-bind GlcNAc isolated from T. rangeli (Tr-60) showed a potent parasite adhesion inhibitory effect indicating that this protein may act as a ligand receptor in the interaction between the trypanosome and the gland cell surface. The presence of SDS (sodium dodecylsulphate) in the buffer did not affect the interaction of this glycosylated protein with the salivary gland cells indicating that the Tr-60 could be a glycoprotein. The purity was confirmed by reverse phase HPLC. REFERENCES
Copyright 2004 -Medical Journal of the Islamic Republic of Iran The following images related to this document are available:Photo images[mr04030f3.jpg] [mr04030f2.jpg] [mr04030f1.jpg] [mr04030t2.jpg] [mr04030t1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}