 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Indian Journal of Medical Science Vol. 58 No. 3, March 2004 , pp. 122-130 EVALUATION OF THE ROLE OF MELATONIN IN FORMALIN-INDUCED PAIN RESPONSE IN MICE M RAY, P K MEDIRATTA, P MAHAJAN, K K SHARMA Department of Pharmacology, University College of Medical Sciences & G.T.B.
Hospital, Delhi - 110 095
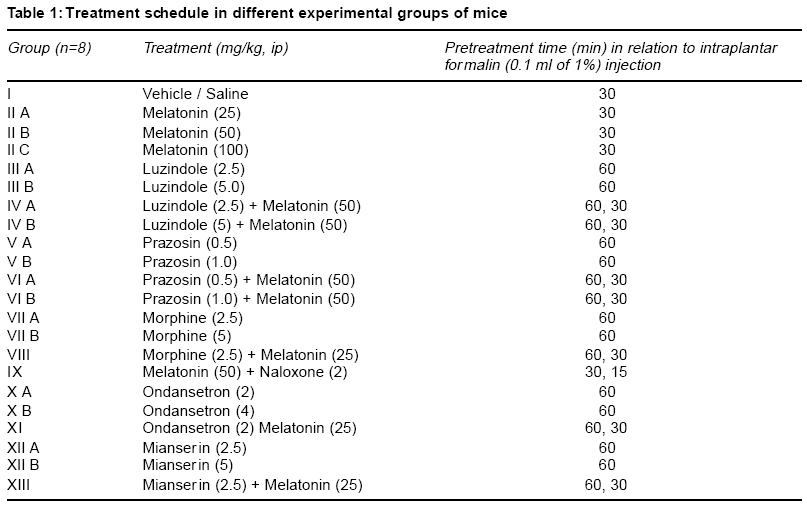
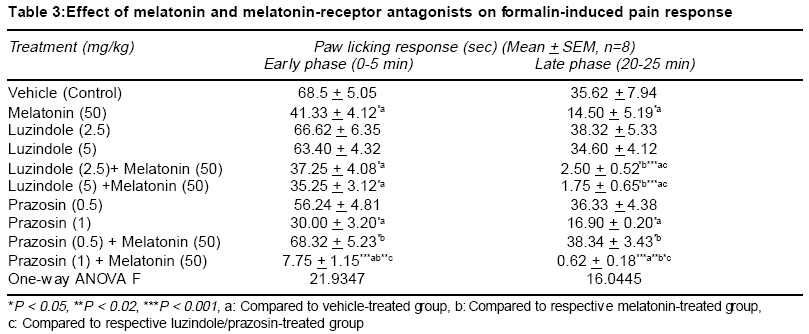
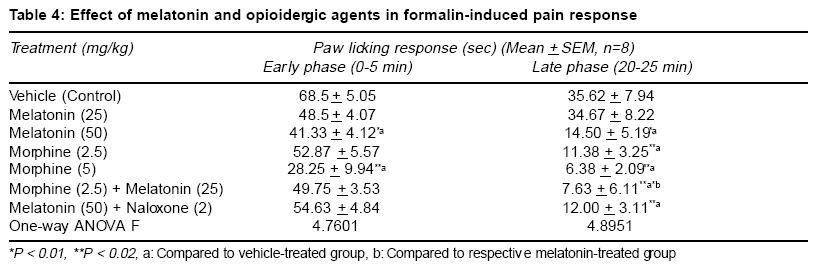
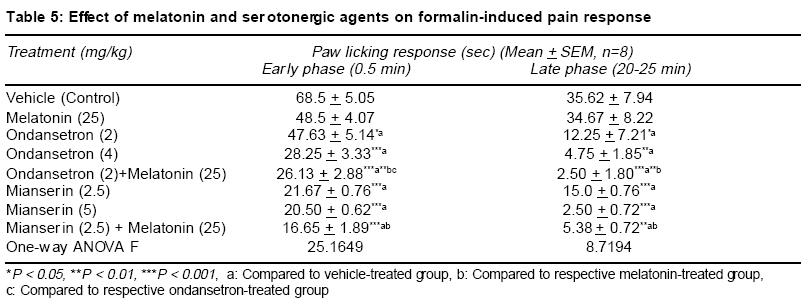
Accepted Date: 22-03-2004 Code Number: ms04019 Abstract Background: Melatonin, the major secretory product of pineal gland has been suggested to play a regulatory role in the circadian rhythm of body activities including the pain sensitivity. Three subtypes of melatonin receptors, i.e. ML1, ML2 and ML3 have been identified. AIM: To investigate the antinociceptive activity of melatonin and to unravel the underlying receptor mechanisms involved in this action. MATERIAL AND METHODS: Effect of melatonin (25-100 mg/kg, ip) and its interaction with putative melatonin receptor antagonists and opioidergic and serotoninergic agents have been studied in formalin test, a model of tonic continuous pain. Formalin (0.1 ml of 1% solution) was injected under the plantar surface of right hind paw of mice and the time an animal spent in licking the injected paw was measured. STATISTICAL ANALYSIS: The data were analysed by one-way ANOVA followed by Tukey's test for multiple comparisons. RESULTS: Injection of formalin produced two phases of intense licking, an early phase (0-5 min) and a late phase (20-25 min). Melatonin dose-dependently decreased the licking response in both the phases, effect being more marked in the late phase. Luzindole, a ML1 receptor antagonist did not block but rather enhanced the antinociceptive activity of melatonin. However, prazosin, a ML2 receptor antagonist in the low dose (0.5 mg/kg) significantly attenuated but in higher dose (1 mg/kg) enhanced the analgesic effect of melatonin. Naloxone, an opioid receptor antagonist did not reverse but morphine, an opioid agonist enhanced the antinociceptive activity of melatonin. Both mianserin and ondansetron the 5HT2 and 5HT3 receptor antagonists, respectively increased the analgesic effect of melatonin. CONCLUSION: The present results suggest the involvement of ML2 receptors in mediating the antinociceptive activity of melatonin in formalin-induced pain response. Further an interplay between melatonin, alpha-1 adrenergic and 5HT2 and 5HT3 serotoninergic receptors may also be participating in this action. Key Words: Melatonin, formalin test, antinociceptive activity, melatonin receptor antagonists, opioidergic agents, serotoninergic agents. INTRODUCTION Melatonin, the major secretory product of pineal gland has been suggested to be a strong synchronizer of a broad spectrum of activities ranging from alteration of skin colour to resetting the circadian clock in mammals, thermoregulation, communication of time of day and season between foetus and mother, regulation of appetite and change in pelage to suit altered environmental conditions.1,2 Three subtypes of melatonin receptors have been identified, i.e. ML1, ML2 and ML3. ML3 receptor is found only in lower vertebrates and birds while ML1 and ML2 are found in higher vertebrates and humans. Melatonin binds very strongly to ML1 receptor, also known as "high affinity" melatonin receptor having slow kinetics of association and dissociation. Binding to ML2 receptor is regulated by temperature. It is the "Low affinity" melatonin receptor and has fast kinetics of association and dissociation3,4. Both ML1 and ML2 receptors have been shown to belong to the family of G-protein coupled receptors.3,5 Melatonin through binding to calmodulin, may also directly affect Ca2+ signaling by interaction with key target enzymes (e.g. adenylate cyclase and phosphodiesterase) and structural proteins (e.g. microtubules and tubuline) and indirectly via specific protein kinase.3-6 Sensitivity and response to pain and melatonin synthesis by the pineal gland have been observed to vary in a circadian manner. Both the melatonin synthesis in the pineal gland as well as its plasma levels were found to be greatest during darkness, at which time animals were reported to be least sensitive to pain. Pinealectomy in rats has been shown to change the levels and circadian rhythmicity of met-enkephalin and b-endorphin, the two endogenous opioids involved in the regulation of pain sensitivity in hypothalamus.7,8 This shows that the change in the brain concentration of these endogenous opioids may be a mechanism for the mediation of the melatonin-induced modulation of pain sensitivity. In conformation to its role in the regulation of pain sensitivity melatonin has been shown to exhibit antinociceptive effect in acute pain due to thermal noxious stimuli.9 So far role of melatonin and its mechanism of action in inflammatory tonic continuous pain response is not well studied. The present study was therefore undertaken to investigate the effect of melatonin in formalin test, a model for tonic continuous pain. Attempt has also been made to find out the underlying receptor mechanisms involved. Both opioidergic and 5HT systems have been reported to be involved in the mediation of pain mechanisms.10,11 Further a link has been shown to exist between melatonin and opioidergic system2,8 and serotonergic system in mediation of other behavioural effect of melatonin.2,12,13 In view of the involvement of these systems in melatonin induced effects, an interaction of melatonin with opioidergic and serotoninergic agents have been studied in order to delineate the role of these neurotransmitter mechanisms in the antinociceptive activity of melatonin. MATERIAL AND METHODS Male albino mice of Swiss strain (25-30 g), obtained from the Central Animal House of the University College of Medical Sciences & G.T.B. Hospital, were used. Each treatment group consisted of 8 animals/group, which were housed groupwise in polypropylene cages (17" x 11" x 6") and kept under controlled environmental conditions (temperature: 22 + 20C, humidity: 50-55%, natural light/dark cycle). The mice received a standard pellet diet (Golden Feeds Ltd., Delhi) and water ad libitum. The animals were allowed to acclimatize in the local laboratory for 1 week prior to the experimentation. All experiments were performed during the light phase between 9.30 AM to 3.30 PM. The protocol of the study was approved by the Institutional Animal Ethics Committee. The care of the animals was as per the Guidelines of Indian National Science Academy, Delhi for the Care and Use of Laboratory Animals. Melatonin (Dabur, New Delhi), prazosin (Sun Pharma, Baroda), ondansetron (Cipla, Mumbai), mianserin, naloxone and luzindole (Sigma, USA) were used in the study. Melatonin was dispersed using 2 drops of Tween 80 and diluted with distilled water. Mianserin, luzindole and naloxone were dissolved in distilled water. Commercially available ondansetron injection was diluted with 0.9% saline. Time interval for injecting various drugs to different groups of animals and their doses were determined by pilot experiments for analgesic activity and on the basis of related studies available in the literature for the maximum biological effects of these agents. Melatonin was administered 30 min, ondansetron, mianserin, luzindole, prazosin and morphine 60 min and naloxone 15 min before formalin injection. Drugs and vehicles were injected in a volume of 10 ml/kg by intraperitoneal (ip) route (Table 1). One percent formalin (0.1 ml) was injected under the plantar surface of the right hind paw of the mouse. The left paw which acted as control was injected with 0.1 ml of 0.9% saline. The mouse was then kept in an open cage and the time spent by the animal licking the injected paw or leg was recorded for a period of 30 min. Two phases of intense licking were observed, an early first phase from 0 5 min and a late second phase between 20 25 min. The first phase represents the acute pain response while the second phase, tonic pain response.14,15 These two phases were scored separately for studying the effect of drugs on formalin-induced pain responses. The data are expressed as mean + SEM. The comparisons between various groups were obtained by one way ANOVA followed by Tukey's test. Differences in p < 0.05 between experimental groups at each point were considered statistically significant. RESULTS In the control animals, the licking time was 68.5 + 5.05 sec (mean + SEM) in the early phase and 35.62 + 7.94 sec during the late phase (Table 2). Melatonin (25-100 mg/kg, ip) dose-dependently decreased the licking response in both early and late phases, the analgesic effect being significant with melatonin in doses of 50 and 100 mg/kg (p<0.01 and p<0.001, respectively). This effect was more pronounced on the late phase as compared to early phase (Table 2). Luzindole (2.5 5 mg/kg, ip), a ML1 receptor antagonist, per se did not modulate the licking response both in the early and late phases. Administration of luzindole with melatonin (50 mg/kg) did not attenuate the analgesic activity of melatonin, rather there was an enhancement of the antinociceptive effect of melatonin, the difference being significant (p<0.05) in the late phase (Table 3). Unlike luzindole, prazosin (0.5 - 1 mg/kg, ip), a ML2 receptor antagonist, per se produced a dose dependent decrease in licking response in both early and late phases, the effect being significant (p<0.05) with the higher dose only. Pretreatment with lower dose of prazosin (0.5 mg/kg) produced a significant reversal of the analgesic effect of melatonin. However, higher dose of prazosin (1 mg/kg) did not reverse the antinociceptive effect of melatonin (50 mg/kg); rather there was a significant increase in the analgesic effect of melatonin, both in the early (p<0.001) and late phases (p<0.02) (Table 3). Morphine (2.5 5 mg/kg, ip) dose-dependently produced a decrease in licking response, both in the early (p < 0.01) and the late phase (p<0.02). Administration of a lower dose of morphine (2.5 mg/kg) along with melatonin (25 mg/kg) enhanced the antinociceptive effect of the latter drug which was significant (p<0.01) only in the late phase (Table 4). Naloxone (2 mg/kg, ip) when administered to melatonin (50 mg/kg) treated animals, failed to reverse the analgesic effect of melatonin both in the early as well as the late phase (Table 4). Injection of ondansetron (2-4 mg/kg, ip) dose-dependently produced a decrease in licking response, both in early (p<0.02; p<0.001) and the late phase (p<0.05; p<0.01) as compared to control group (Table 4). Ondansetron (2 mg/kg) administration with melatonin (25 mg/kg) significantly enhanced the analgesic response produced by the latter both in the early (p<0.01) as well as in the late phase (p<0.001). Like ondansetron, mianserin (2.5 5 mg/kg, ip) also produced a dose-dependent decrease in licking response, both in the early and late phases (p<0.001) and injection of mianserin (2.5 mg/kg) along with melatonin (25 mg/kg) enhanced the analgesic effect produced by melatonin alone, which was significant both in the early (p<0.001) as well as in the late phase (p<0.01) (Table 5). DISCUSSION The results of the present study show that melatonin (25-100 mg/kg) dose-dependently produced an anti-nociceptive effect in formalin-induced tonic pain model. The effect was more marked on the late phase as compared to early phase of pain response. Higher doses (>100 mg/kg) of melatonin were not used as these doses produced motor incoordination. The late phase of licking response in formalin test could be analogous to an acute inflammatory process. There is increased vascular permeability and leukocyte migration occurs. Several mediators along with neutrophil-derived reactive oxygen species and free radicals like peroxynitrite are reported to be involved during this process.16 The greater effectiveness of melatonin in the late phase could be attributed to its ability to inhibit peroxynitrite formation and poly ADP-ribose synthetase activation.6,17,18 The melatonin induced antinociceptive effect was antagonized by a lower dose of prazosin, a putative ML2 receptor antagonist but not by luzindole, a ML1 receptor antagonist. Rather there was an enhancement of melatonin- induced antinociceptive response with luzindole in the late phase. Besides, prazosin in a higher dose was also observed to enhance the antinociceptive effect of melatonin in both early and the late phases of formalin-induced pain paradigm. The reason why luzindole enhanced the analgesic effect of melatonin in the late phase of formalin test could be due to the blockade of ML1 receptors by this compound which would enhance the action of melatonin on ML2 receptors, resulting into an increase in the melatonin-induced antinociception. This suggestion also explains the observation that prazosin, a putative ML2 receptor antagonist in low doses (0.5 mg/kg) was observed to antagonize the analgesic effect of melatonin. However, when given in a higher dose (1 mg/kg), alpha1-adrenoceptor blocking activity of prazosin is perhaps playing a more important role in its interaction with melatonin. It has been reported that thermal hyperalgesia caused by chronic constriction of the sciatic nerve was relieved by tizanidine, an alpha-2 adrenergic receptor agonist.19 There are other reports which indicate that alpha2-adrenergic receptors play an important role in mediation of analgesic responses.20 Blockade of alpha1- adrenergic receptors by prazosin (in a higher dose) would allow the spontaneous released noradrenaline in the CNS to act on alpha2-adrenoceptors, thus leading to enhancement of the antinociceptive activity of melatonin. The antinociceptive activity of melatonin in the late phase was significantly enhanced by morphine, an opioid agonist; however, naloxone, an opioid antagonist failed to modulate the analgesic effect of melatonin. The enhancement of antinociceptive action of melatonin by morphine, may make the central opioid system one of the candidates for mediation of decreased pain response to this indole amine. However, the observation that the analgesic activity of melatonin could not be blunted either in the early or late phase of formalin-induced nociception by naloxone, an opioid antagonist, deny such a mediation of opioid system in the analgesic effect of melatonin. Contrary to our observations, an earlier study has reported reversal of analgesic effect of melatonin by naloxone.9 However these workers have used hot plate test which is a model of acute pain. The formalin-induced pain response is a model of tonic nociceptive activity14,15. Hence, the difference in our observation compared to other workers could be related to the use of different models of nociception, which may be processed differently in the CNS. The observed enhancement of melatonin-induced analgesia with morphine in the present study could be because morphine is reported to exert its antinociceptive effects, in part by enhancing the release of endorphins and enkephalins into the CSF from synaptic clefts and intracellular pools.21 Pinealectomy or melatonin injection has been shown to cause modification in brain [met5] enkephalin and b-endorphin levels.7,8 Thus morphine may be enhancing the analgesic effect of melatonin by augmenting the release of some endopioids, like leu- and met-enkephalins or some other endorphins. As regards the interaction of melatonin with serotoninergic agents, in the present study the 5HT3 antagonist ondansetron per se exhibited an analgesic response in formalin test in both early and late phases. Further, the analgesic effect of melatonin was enhanced by ondansetron in both phases. Similar results were also observed with the 5HT2 antagonist, mianserin. The midbrain periaqueductal gray is believed to be an important component in the endogenous pain control system and it has been suggested that 5HT modulates sensitivity of pain by acting through different 5HT receptors on neurons conducting pain pathways. The antinociceptive effect of 5HT3 antagonists could be hypothesized from the presence of a dense band of 5HT3 receptors on capsaicin sensitive primary afferent terminals in the superficial dorsal horn of the spinal cord22. Stimulation of 5HT3 receptors by 2-methyl 5HT has been shown to block scratching and biting responses induced by substance P and NMDA, applied intrathecally in mice. A link between melatonin and the central serotonergic system has been proposed to exist.12,13 Intraperitoneal administration of melatonin has been shown to produce an elevation in brain serotonin levels. Besides, serotonin was also observed to antagonize melatonin-induced behavioural changes after injection into nucleus accumbens of rats12. It has been suggested that coactivation of 5HT2 and 5HT3 receptors located on capsaicin-sensitive afferents may be mediating the nociceptive response.22 This may explain the analgesic activity observed in the present study with mianserin and ondansetron alone and in combination with melatonin. Results of the present study thus show that melatonin by acting on ML2 receptors exerts an antinociceptive effect on formalin-induced continuous pain response and a number of mechanisms including modulation of levels of enkephalins and endorphins may be involved. There appears to be an interplay between various receptors like melatonin receptors alpha1, adrenergic receptors and 5HT2 and 5HT3 serotoninergic receptors. Further, its action on the late phase can also be attributed to decreased production of reactive oxygen species and other free radicals like peroxynitrite. These results further suggests the potential of melatonin as an adjunct therapy during pain specially when an inflammatory component is involved. REFERENCES
Copyright by The Indian Journal of Medical Sciences The following images related to this document are available:Photo images[ms04019t4.jpg] [ms04019t2.jpg] [ms04019t5.jpg] [ms04019t3.jpg] [ms04019t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}