 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Indian Journal of Medical Science Vol. 58 No. 4, April 2004 , pp. 155-160 Effect of undernutrition and subsequent rehabilitation on brain 5-HT (serotonin) profile of developing newborn rats K P Singh, A K Sanyal* Clinical Research Unit (H), Centre of Experimental Medicine and
Surgery, and *Department of Pharmacology, Institute of Medical Sciences, Banaras
Hindu University, Varanasi - 221005, India.
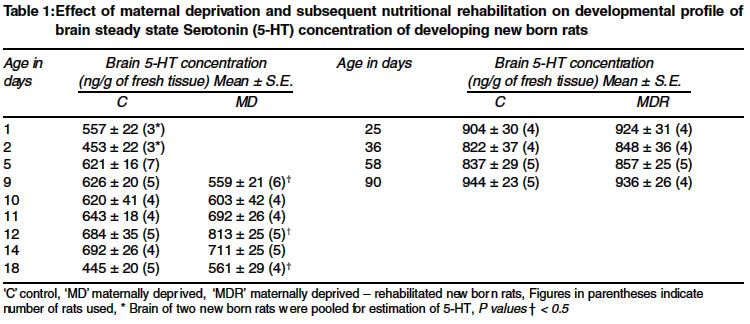
Code Number: ms04026 ABSTRACT For littermate comparison, undernutrition in half of the pups of a litter was induced by 12h maternal deprivation from day 5 to day 18 postnatal. Subsequently, undernourished suckling rats were rehabilitated. Brain 5-HT concentrations and its turnover rate were measured on different days of age up to the period of 3 months. Brain 5-HT concentration was approximately half of the adult level at birth and increased progressively, except with a steep fall on day 18 postnatal, to the adult level by day 25 postnatal and thereafter it did not vary much on different days of age. Undernourished newborn rats showed significantly low level of brain 5-HT on day 9 and significantly higher concentrations on day 12 and 18 postnatal, whereas, rehabilitated rats showed comparable brain 5-HT concentrations with that of their nourished littermates. It is concluded that undernutrition had affected the brain 5-HT concentration and its turnover rate. Brain 5-HT concentration was low during early phase and was more during late phase of undernutrition. Rehabilitation of undernourished rats restored the brain 5-HT concentration similar to that of their nourished littermates. Key Words: Nutrition, Serotonin, Animal experiment. INRODUCTION The existing literature on the effect of undernutrition on brain 5-HT (serotonin) steady state levels and its turn over rate is full of controversy. Some workers have reported increased levels1 while others have reported a decreased levels2,3 or no change in its concentrations.4 Such discrepancy in brain 5-HT levels of undernourished newborn rats could be either due to the variations in the duration of nutrition insult imposed or to the different methods used for inducing undernutrition by these workers. In these studies,1-4 interlitter nutritional status of both nourished and undernourished groups varied widely as the lactating mothers rearing the different litters might have demonstrated differential lactating capacity or there was an intralitter mutual competition for the teats in which relatively stronger pups commonly displaced the weaker ones. The above premise, therefore, prompted us to study the effect of undernutrition in the pups of a same litter in which 50% of the pups of a litter were undernourished by 12h of maternal deprivation. MATERIAL AND METHODS Induction of malnutrition in new born rats Undernutrition in developing newborn rats was induced by 12h (from 9.00 pm to 9.00 am next morning) maternal deprivation of 50% of the pups of a litter according to the method of Mishra et al5 as described earlier.6 Thereafter, undernourished pups were rehabilitated on normal feeding similar to that of nourished littermates. Estimation of brain serotonin and rate of accumulation Brain 5-HT concentration (whole brain minus cerebellum) was estimated according to the method described by Curzon et al.7 In short, new born rats were decapitated between 9.30-10.30 A.M. Brain was quickly dissected out and put in 6-8 ml ice cooled acid-butanol. After removing both cerebelli, the brain was weighed and homogenized for 3 min in a Potter Elvehjem homogenizer with 10 volumes of ice cooled acid-butanol. The content was centrifuged at 3000 rpm for 5 min and the supernatant (4 ml) was pipetted into glass stoppered tube containing 8 ml of ice cooled n-heptane and 0.6 ml of 0.1 N. HCl (containing Lcysteine 1mg/ml). The mixture was shaken for 5 min on a wrist action shaker and centrifuged again for 5 min. From it, 0.3 ml of aqueous phase was transferred into 12 x 150 mm glass stoppered tube. After adding 2 ml of conc. HCl (containing Lcysteine), the content was mixed for 3 min on vortex shaker. Afterwards, the tubes were heated in a boiling water bath for 15 min and then cooled in running water. The content was transferred to a quartz cuvette and the fluorescence was measured at an emission wave length of 470 nm after excitation at 360 nm in an Aminco Bowman spectro-photofluorometer against a reagent blank and working standard. The concentration of brain 5-HT has been expressed as ng/g of fresh tissue. The turn over rate of 5-HT in brain was estimated according
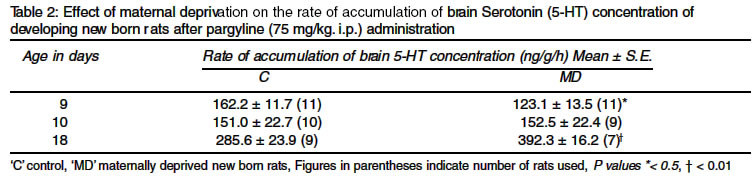
to the method of Neff et al8 by measuring the accumulation of brain RESULTS Effect of undernutrition and subsequent rehabilitation on developmental profile of brain 5-HT The mean 5-HT levels of whole brain (excluding cerebellum) in between 12-16h of the birth, was 557 ± 32 ng/g of fresh tissue. On the second day of life its level decreased significantly (453 ± 22 ng/g). Subsequent estimation from day 5 to day 14 postnatal demonstrated a gradual increase in brain 5-HT levels. Surprisingly, there was a steep fall in brain 5-HT levels (445 ± 20 ng/g) on day 18 postnatal. It rose again on day 25 postnatal (904 ± 30 ng/g) followed by a slight fall on day 36 (Table 1). Thereafter it rose very slowly to attain brain 5-HT levels similar to or slightly higher than that observed on day 25 postnatal. The mean 5HT level estimated on day 90 postnatal was the order of 944 ± 23 ng/g of fresh brain tissue. The developmental profile of brain 5-HT in undernourished developing newborn rats was qualitatively identical but quantitatively distinctly different to that of their respective nourished littermates. Initially, on day 9 postnatal, undernourished newborn rats showed significantly lower brain 5-HT concentration (559 ± 21 ng/g) as compared to that of nourished littermates (626 ± 20 ng/g). Afterwards, brain 5-HT concentrations in undernourished animals increased at a much faster rate, which were significantly higher on day 12 and day 18 postnatal. Undernourished newborn rats rehabilitated for one week or more showed comparable brain 5-HT levels to that of their respective nourished littermates (Table 1). Effect of undernutrition on the rate of accumulation of brain 5-HT Accumulation of brain 5-HT was significantly low on day 9 (123.1 ± 13.5 ng/g/h as compared to 162.2 ± 41.7 ng/g/h), practically same on day 10 (152.5 ± 22.4 ng/g/h and 151.0 ± 22.7 ng/g/h) and significantly higher on day 18 postnatal (392.2 ± 16.2 ng/g/h as compared to 285.6 ± 23.9 ng/g/h) in undernourished newborn rats when compared with nourished littermates (Table 2). DISCUSSION Available literature on the normal developmental profile of brain 5-HT2,9 and the effect of undernutrition1-4 on its steady state level and turn over rate in developing newborn rats is equivocal. In spite of some disagreement, there is a general consensus that brain 5-HT level is low at birth and gradually increases to attain the adult levels by 5-6 weeks of age.3,9,10 In the present study, brain 5-HT levels when measured between 12-16 h after birth on day 1 was 557 ± 22 ng/g of fresh tissue. It's level decreased by 19.6% on day 2 and then progressively increased, except a steep fall on day 18 postnatal, to attain the adult levels by day 25 postnatal. Thereafter, brain 5-HT levels remained `more or less' steady till the last day of observation i.e. on day 90 postnatal. Higher levels of brain 5-HT immediately after birth has also been reported by Bourgoin et al.10 An increase in brain 5-HT level has been reported by Sanyal et al11 after 1-2 h of immobilization stress and by Vermes et al12 after ACTH and corticosterone administration. Holt et al13 have reported 3.5 fold increase in plasma corticosterone levels within 5h of birth and suggested that stress of parturition was responsible for higher rate of corticosterone production by rat's adrenals. It is presumed, therefore, that the higher level of brain 5-HT observed in the present study within 16h of birth could be due to the release of glucocorticoids12 associated with the stress of parturition,13 while, lower levels of 5-HT in brain observed on the second day of birth could be due to passing of the stress associated with the parturition. The observed low level of brain 5-HT at birth could be related to the poor development of serotonergic innervation in brain.9,10 Progressive increase in brain 5-HT from day 2 onward to day 14 postnatal might be due to simultaneous development of serotonergic innervation and increase in tryptophan hydroxylase activity.14 Likewise, the steep fall in brain 5-HT levels observed on day 18 postnatal could be explained on the basis of its rapid catabolism since the rate of accumulation of brain 5-HT was maximum (285.6 ± 23.9 ng/g/h) on this day as compared to 9 days old new born developing rats (162.2 ± 11.7 ng/g/h) and matured adult rats (216.6 ± 13.4 ng/g/h) respectively, after inhibition of MAO by pargyline. Bourgoin et al15 had also reported 80% higher MAO activity in 15 days old rats as compared to that of adult ones. The development of serotonergic innervation and tryptophan hydroxylase activity to its maximum levels14 and decreased MAO activity15 in matured rats could be the reason for the higher values of brain 5-HT observed in the present study in 25 days old rats. Afterwards, brain 5-HT levels of adult rats maintained on a balanced diet were `more or less' steady when estimated on different days of age. Likewise undernourished developing newborn rats showed wide variations in brain 5-HT concentration when estimated on different days of their life. It's level decreased significantly on day 9 postnatal i.e. just after 3 days of the initiation of the undernourishment (559 ± 21 ng/g as compared to 626 ± 20 ng/g in nourished littermates), became more or less same on day 10 and 11 and were significantly higher on day 12 (813 ± 25 ng/g as compared to 648 ± 35 ng/g in nourished littermates) and on day 18 (561 ± 29 ng/g as compared to 445 ± 20 ng/g in nourished littermates) but not on day 14. When 18 days old undernourished developing rats were rehabilitated on normal feeding, they showed comparable brain 5-HT concentrations to that of their nourished littermates. If rate of accumulation of 5-HT is considered to be an index of the rate of synthesis,8,11 then the rate of synthesis of 5-HT was found to be significantly less on day 9, equal on day 10 and significantly more on day 18 postnatal in undernourished newborn rats as compared to that of their nourished littermates, implying that serotonergic neuronal activity of undernourished brain of newborn developing rats was considerably less during the early phase of undernourishment but was significantly more during the established phase of undernutrition. On the other hand, rehabilitated undernourished developing rats showed comparable brain serotonergic neuronal activity with that of their respective nourished litterm ates. ACKNOWLEDGEMENTS Author (KPS) is grateful to Prof. K. D. Gode, Prof. S. K. Bhattacharya, Prof. S. S. Gambhir, Prof. R. K. Goel of Pharmacology Department and Prof. Raj Shankar of Biochemistry Department, Institute of Medical Sciences, Banaras Hindu University, Varanasi, for their technical guidance and help rendered during this work. REFERENCES
Copyright by The Indian Journal of Medical Sciences The following images related to this document are available:Photo images[ms04026t2.jpg] [ms04026t1.jpg] |
| |||||||||

{kind=link}
{kind=link}