 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
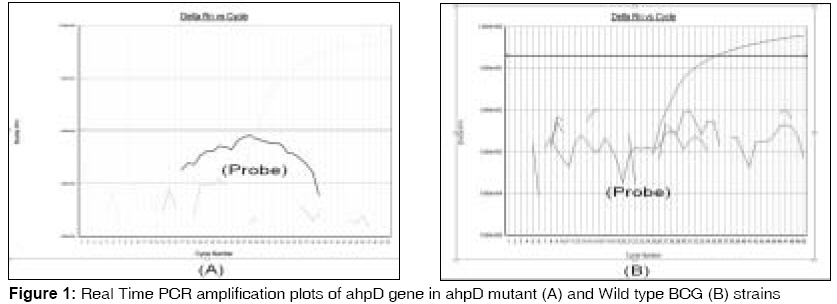
Indian Journal of Medical Sciences, Vol. 62, No. 7, July, 2008, pp. 275-282 ORIGINAL CONTRIBUTION Mutation in alkylhydroperoxidase D gene dramatically decreases persistence of Mycobacterium bovis bacillus calmette-guerin in infected macrophage Farivar Taghi Naserpour, Varnousfaderani Pouran Johari, Borji Abasalt School of Medicine, Research Institute for Tropical Medicine and Infectious Diseases, Zahedan University of Medical Sciences, Zahedan Code Number: ms08049 Abstract Background and Objectives: Mycobacterium tuberculosis is the leading cause of death from a single bacterial species in the world and is subjected to a highly oxidative environment in its host macrophage and consequently has evolved protective mechanisms against reactive oxygen and nitrogen intermediates. Alkyl hydroperoxidase D (AhpD) is a molecule from these mycobacterial defense systems that has a dual function. It not only works with Alkyl hydroperoxidase C (AhpC) in mycobacterial defense system against oxidative stress but also has a role in oxidation/reduction of succinyltransferase B (SucB), dihydrolipoamide dehydrogenase (LPD) and AhpC. The present study was undertaken to find out the effects of inactivation of ahpD gene in the intra-macrophage persistence of resulted BCG mutant.Materials and Methods: We did allelic exchange mutagenesis in Mycobacterium bovis BCG and evaluate the effects of this mutagenesis in intracellular persistence of wild type BCG strains and ahpD mutant ones by comparing colony forming units (CFU) in infected macrophage. Results: Our findings showed that after producing allelic exchange mutagenesis in ahpD gene of M.bovis BCG a sever decrease in the CFU's of ahpD mutant BCG strains has been observed and intracellular persistence of ahpD mutant BCG strains decreased significantly. Conclusion: Mutagenesis in ahpD gene will cause significant decrease in intracellular survival of ahpD mutant strains than wild type M.bovis BCG strains and could leads to an inefficiency in pyruvate dehydrogenase pathway and could also impair impairs mycobacterial defense system against oxidative and nitrosative stress. Keywords: AhpD, macrophage, mycobacteria, Mycobacterium bovis, persistence Introduction In most infected individuals, oxidative and nitrosative stress controls proliferation of Mycobacterium tuberculosis in infected macrophage [1],[2] and in these host macrophages, M. tuberculosis is subjected to a highly oxidative environment. [3] In this situation, produced peroxynitrite (ONOO-) and other reactive nitrogen and oxygen intermediates plays an important role in host defense against the invading bacteria. [4],[5] Consequently, many bacterial pathogens have evolved protective mechanisms against reactive oxygen and nitrogen intermediates. [6] On the other hand, it has been cleared that mutation in the catalase-peroxidase katG gene of M. tuberculosis resulted in resistance to isoniazid [7] and mutation in the flavoprotein mono-oxygenase etaA gene, resulted in resistance to ethionamide. [8] Interestingly, although KatG and EtaA have different activating enzymes, mutations of katG and etaA , resulted in elevated expression of alkyl hydroperoxidase C (AhpC) [7] and analysis of the genes induced in isoniazid-resistant M. tuberculosis strains indicated that up-regulation of ahpC gene, is one of the mechanisms used by the organism to restore the loss of the KatG protein antioxidant activity. [9] AhpC is a member of the ubiquitous peroxiredoxin family and its disulfide bond is reduced by different mechanisms to give a sulfenic acid (-SOH) intermediate in different organisms. [10] In M. tuberculosis thioredoxin and thioredoxin reductase, do not reduce the corresponding AhpC. Previous studies showed that an alkyl hydroperoxidase D ( ahpD ) gene (Rv2429) coding for AhpD protein with no sequence uniqueness to AhpC is located instantly next to the ahpC gene and it works as reducing partner of AhpC. AhpD is reduced by dihydrolipoamide succinyltransferase (SucB) and dihydrolipoamide dehydrogenase (LPD). [11] AhpD is a homo trimer and contains two cysteines which are required for enzymatic activity of AhpD protein. [12] Thus, AhpC, AhpD, SucB, and Lpd together constitute a peroxidase active toward both hydrogen and alkyl peroxides. [13] These studies have yielded a potent in vitro inhibitor of AhpD that has been used to explore the potential of AhpD as a target for antituberculosis drug development. [14] These findings showed that AhpD has a dual role in metabolic pathway of pyruvate dehydrogenase and in anti-oxidative pathway of mycobacteria. As low titer of AhpD suffices to maintain AhpC activity and known inhibitors of AhpD do not completely suppress the in vitro activity of AhpC/AhpD, using competitive precursor for AhpD could not inactivate the AhpC/AhpD complex. [14] So, we did this study to determine the effects of allelic exchange mutagenesis in ahpD gene on the persistence of Mycobacterium bovis Bacillus Calmette-Guerin(BCG) in infected Macrophages. Materials and Methods Materials Media and growth conditions Cloning and allelic exchange mutagenesis The reaction contained 50 ng of M. bovis BCG Pasteur strain 1173P2 genomic DNA, 50 pmol each of the primers, 2 mM dNTPs, and 10 units of Vent DNA polymerase in a final volume of 100 µl of 20 mM Tris-HCl (pH 8.8), 10 mM KCl, 10 mM (NH4)2SO4, 2% dimethyl sulfoxide, and 0.1% Triton X-100. The annealing and extension cycles were as follows: 90 °C for 10 min (1 cycle), 90 °C for 1 min, 72 °C for 1 min, 60 °C for 1 min (30 cycles), and 72 °C for 10 min (1 cycle). [15] Disruption of ahpD gene: we disrupted ahpD gene in M. bovis BCG Pasteur strain 1173P2 by using the gene replacement and transposon delivery strategy by Guilhot [16] method. Briefly, following gel purification of the amplified ahpD gene, 0.55-kilobase product gene was digested with PstI and the ligated two resulted fragments with hygromycine resistance cassette resulted from digestion of pGoal 19 with pstI. Then we cloned the ligated fragment into pPR23( ts-sac B vector) with cutting this plasmid by KspAI (pPR23ahpD plasmid). Resulted plasmid electroporated into M. bovis BCG Pasteur strain 1173P2 by gene pulser(Bio-Rad,Tehran,Iran) with the following conditions; Single Pulse; 2.5 KV;1000 Ohms;25µF [16] and then we selected mutant BCG by culturing them on hygromycin containing media which had 2% sucrose. After culturing mutant strains in medium A for 14 days at 37°C, 2 ml of liquid medium was selected and after centrifugation at 8000 rpm for 10 min, total RNA of isolated bacteria was purified by Max kit (Ambion, Austin, USA) and underwent for Real Time PCR. Real time PCR Real-time PCR was performed using an ABI Prism 7500 System (Applied Biosystems, Foster City, CA).Each 50-µl reaction mixture consisted of 40 µl of PCR mix combined with 10 µl of internal standard, control, or specimen DNA. The PCR mix contained universal master mix (Applied Biosystems), primers forward and reverse (each at 50 nmol/reaction), probe (10 nmol/reaction), and 13 µl of RNase/DNase-free water. After 2 min at 50°C and 10 min at 95°C, there were 45 cycles (95°C for 15 s and 60°C for 1 min) of PCR amplification for AhpD gene detection. AmpErase and dUTP within the master mix provided carryover contamination control. Bacterial cell association and replication Human peripheral blood mononuclear cells Cell suspensions of the M. bovis BCG Pasteur 1173P2 and mutant BCG were added to the attached macrophages at a multiplicity of infection (MOI) of 1:10 (1 bacterium per 10 host cells). Each day, the infected macrophages were washed twice with Hank′s Balannce Salt Solution (HBSS) and overlaid with fresh Iscove′s Modified Dulbecco′s Medium (IMDM). [17] CFU assay: Adherent monolayers were disrupted with a solution of water containing 0.016% Digitonin and 0.25% Tween 80 (Sigma Chemical Co.). Bacterial suspensions were serially diluted and plated onto Middlebrook 7H10 agar plates supplemented with oleic acid-albumin-dextrose-catalase enrichment (Difco). Plates were incubated for three hours, one day, eight days and 15 days at 37°C. Colonies were counted under a dissecting microscope and reported as CFU. For each culture dilution, six replicate samples were plated and the mean number of colonies was calculated. [20] Stimulation of Growth of ahpD mutant: Stimulation of growth of ahpD mutant was done by adding the following substances: Thiamine 150 µg/ml, Thymine 8 µg/ml, Guanine 45 µg/ml, Adenine 920 µg/ml, Sodium acetate 3.8 mg/ml, Succinic acid 5 mg/ml. [21] Almar Blue Results Mutagenesis in ahpD gene was confirmed by Real Time PCR of the Wild and ahpD mutant strain after total RNA extraction from mutant and wild type BCG culture and reverse transcription [Figure - 1]. This mutagenesis resulted to production of ahpD mutant BCG strains which were not able to grow in routine mycobacterial culture media (Middlebrook 7H9 or 7H10) as well as do wild type strains. These mutants were only able to growth in these media in the presence of supplementary materials. So, like previous experiments, [21] we added supplements to culture media to enhance mutant growth rate. We added supplements into our three BCG mutant test groups and maintained one group without adding supplements as control. The maximum growth was observed in group A medium which contained all kinds of supplements mentioned in [Table - 1], followed by the group B, which had not contain succinic acid and sodium acetate and the least growth was observed in group C which had only succinic acid and sodium acetate [Table - 1]. Growth curve of wild and ahpD mutant BCG strains were showed in [Figure - 2]. Intracellular persistence of both wild and mutant BCG strains were analyzed by counting viable CFU after one, eight and 15 days of infection in six different wells for each group and the results are mean of these amounts. As described in [Figure - 3], both strains failed to grow, but they were not killed at similar rates by the macrophages. These results for kinetics of wild type intracellular BCG agree with those from previous studies. [22] Direct observation of the cells with acid fast staining (Kinyoun) demonstrated that the percentage of cells associated with mycobacteria (37%) were the same for wild type and mutants but the number of the two strains during the incubations was different. Taken together, these results show that mutagenesis in ahpD gene significantly affects BCG growth inside infected cultured macrophages. Our study showed that after a transient increase in the CFU of lysates of the infected macrophage by the wild type BCG strains in the first day, the CFU of these lysates decreased in the next days. On the other hand, ahpD mutant strains in addition to the growth problems which mentioned before, were killed rapidly in infected macrophages in such a way that their CFU′s immediately decreased from the first day and in the fifteenth day reached to its least count number. The ratio of CFU′s of ahpD mutant strains to wild type one′s decreased 700 and 45 fold in days one and eight respectively and 60% fold in day fifteen [Figure - 3] and so, there is a significant relation between mutation in ahpD gene and decreasing survival of the resulted mutants. Also Almar Fluorescence test showed 0.03, 2.5 and 60 percent mean decrease in the fluorescent emission of wild than ahpD mutant BCG in first, eight and fifteen days respectively [Figure - 4]. Discussion Our study showed that mutation in ahpD gene make mutant strains susceptible to intra-cellular killing power of infected macrophage. Also, our study showed that mutation in ahpD gene make mutant strains unable to growth in routine mycobacterial culture. We did this study to find whether mutation in ahpD decrease intra-cellular survival of mutant strains. For evaluating this question, we did allelic exchange mutagenesis in ahpD gene of M.bovis BCG and then we infected macrophage culture with wild and mutant BCG strains and evaluated the intra-cellular survival of these strains by colony counting and Almar blue assay in six wells for each group. We compared means of colony count CFU and Almar fluorescent of wells in wild and mutant BCG strains. Analysis of these data showed that there is a significant relation between mutation of ahpD gene and decrease of intra-cellular survival of wild and mutant BCG strains. Our study showed that ahpD not only take part in the oxidative and nitrosative resistance mechanisms of BCG but also, it has a critical role in the growth of BCG strains in routine mycobacterial culture. Also, our findings showed that there is some substitutive mechanism(s) that make mutant BCG strains to survive in infected macrophage although very weak. On the other hand, Koshkin study [14] showed that ahpD is an element of peroxiredoxin defense against oxidative stress. Also Hillas and his colleague [15] reported that the ahpC and ahpD are anti-oxidant defense system of M. tuberculosis . As Bryk and his colleague [13] reported, if we could be able to inhibit SucB or Lpd in M. tuberculosis without affecting their human counterparts, both the Krebs cycle and the bacillus′s ability to synthesize acetyl-coenzyme A (CoA) from endogenous precursors could be in danger and our study suggests that AhpD is a good candidate for this purpose. Acknowledgments We thank Dr. Tanya Parish for pGoal 19 plasmid and Dr. Jean Rauzier for pPR23 and Dr. Nader Shahrokhi for electroporation of the mutants.References
Copyright 2008 - Indian Journal of Medical Sciences The following images related to this document are available:Photo images[ms08049f4.jpg] [ms08049f1.jpg] [ms08049f3.jpg] [ms08049f2.jpg] [ms08049t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}