 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
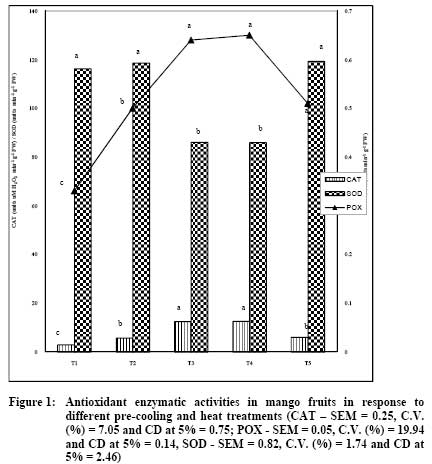
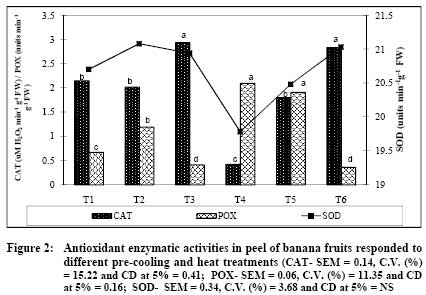
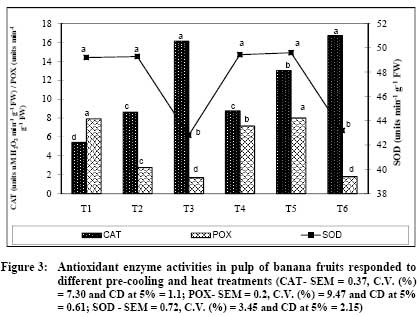
African Journal of Food Agriculture, Nutrition and Development, Vol. 9, No. 5, May, 2009, pp. 1210-1225 Article Effect of pre-cooling and heat treatment on antioxidant enzymes profile of mango and banana Niranjana P, Gopalakrishna KPR, Sudhakar DVR, Madhusudhan B Department of Studies in Biochemistry and Food Technology, Post Graduate Centre, Kuvempu University, Shivagangotri, Davanagere-577 002 Correspondence Address:Department of Studies in Biochemistry and Food Technology, Post Graduate Centre, Kuvempu University, Shivagangotri, Davanagere-577 002 basavaraj_madhu@hotmail.com Code Number: nd09056 Abstract Fruits have self defense mechanisms to protect from oxidative stress by the activation of many antioxidant defense enzymes like superoxide dismutase (SOD), catalase (CAT) and peroxidase (POX). The development of oxidative stress in fruit mainly depends upon its cellular antioxidant levels, physical atmosphere of the fruit and its postharvest handling. The present investigation was carried out to study the effect of post harvest pre-cooling and heat treatments on antioxidant enzymes, CAT, POX and SOD involved in detoxification of reactive oxygen species (ROS) were measured in mango var. Alphonso and banana var. Robusta. Increased CAT and POX activities in mango and banana during heat treatment followed by cooling at 8 ºC or 13 ºC indicated increased elimination of ROS. Pre-cooling of mango at 8 ºC showed two-fold decreased CAT activity in the pulp whereas heat treatment to mango followed by cooling at 8 ºC and 13 ºC recorded 2.2 fold increased CAT activity. Pre-cooling treatment reduced POX activity whereas heat treatment followed by cooling resulted in decreased SOD activity. CAT activity after pre-cooling to 13 ºC by using pre-cooled air was similar to that of control. In banana, pre-cooling treatment (8 ºC) and heat treatment followed by cooling reduced CAT activity in peel and pulp, whereas POX activity increased. Pre-cooling and heat treatments altered normal homeostasis of these fruits, which were reflected by changes in the levels of antioxidant enzymes- CAT, POX and SOD. Visual assessment of mango and banana fruits after pre-cooling at different temperatures (5 ºC, 8 ºC and 13 ºC) did not show any chilling injury (CI) symptoms. Similarly, application of postharvest quarantine heat treatment (52-55 °C) for 5 minutes to mango showed no heat injury (HI) symptoms like skin scalding, damaged lenticels which could be due to very short duration of exposure. Extent of recommendation of pre-cooling temperature and heat treatment of these fruits after pre-cooling at 5 ºC and 8 ºC can be made based on the levels of reactive oxygen species. The present study showed that the level of expression of antioxidant enzymes got affected during pre-cooling (5 ºC and 8 ºC) and quarantine heat treatment (52-55 °C). Therefore, findings of the present study serve as basic information about cellular homeostasis in the mango and banana.Keywords: Antioxidant enzymes, heat treatments, pre-cooling Introduction The postharvest losses of fruits and vegetables are mainly attributed to activity of pests and pathogens carried from farm to storage. Thermal quarantine heat treatment (46-48º C for 60 to 90 min) is being adopted world wide as viable non-chemical control method to kill different stages of fruit fly in infected fruits for the prevention of invasive pests of mango, specifically against Mediterranean fruit fly ( Cevatitis capitata ) and Mexican fruit fly ( Anastrepha ludens ) [1],[2] . Temperature is one of the most important environmental factors that influence the deterioration of harvested fruits and hence its management during various postharvest operations like - pre-cooling, pretreatments and storage plays a major role for extending the shelf-life [3] . Pre-cooling can immediately lower the field heat of commodity following harvest and slow down metabolism and reduce deterioration prior to transport or storage [4],[5] . The reduction in temperature has an added advantage in reducing the production and sensitivity of the produce to ethylene that accelerates ripening and senescence [6] . Pre-cooling of fruits harvested in temperate regions was observed to be highly useful in extending the storage life and maintaining the qualities thereby getting better market price [3] . Similarly, low temperature storage has been one of the most effective methods for maintaining the quality of most of the fruits and vegetables. This method reduces the rate of respiration, ethylene production, ripening, senescence, undesirable metabolic changes and further decay [3] . Pre-cooling and heat treatment may affect normal biochemical pathways resulting in the oxidative stress. Reactive oxygen species (ROS) are produced due to various metabolic activities and the production increases under various stress conditions [7],[8] . Reactive oxygen species are partially reduced forms of molecular oxygen resulting from either the excitation to form singlet oxygen (O 2 ) or the transfer of one, two or three electrons to O 2 to form super oxide (O2 - ), hydrogen peroxide (H 2 O 2 ) and hydroxyl radical (OH - ), respectively. Oxidative stress occurs when the generation of ROS exceeds the capacity of the fruit or plant to maintain cellular redox homeostasis. The generated ROS affect metabolism of the fruit or plant by oxidative damage to various cellular compartments including membrane lipids, proteins and nucleic acids [9] . Fruits have self defense mechanism to protect from oxidative stress by the activation of many antioxidant defense enzymes like SOD, CAT and POX. The development of oxidative stress in the fruits mainly depends upon its cellular antioxidant levels, physical atmosphere of the fruit and its post harvest handling. Alphonso and Robusta of mango and banana respectively, are excellent varieties having high export value in the world market and also being consumed on a larger scale than other varieties. The changes in carbohydrates and texture, carotenoids and ascorbic acid, phenol and other quality attributes during ripening at ambient conditions and low temperature storage/ripening have been well studied [10],[11],[12],[13] . Relatively very meagre work has been carried out on the antioxidant enzymes (CAT, POX and SOD) during pre-cooling and thermal quarantine heat treatments. Therefore, presently efforts have been concentrated to study the responses of the antioxidant enzymes during postharvest pre-cooling and heat treatments of mango variety Alphonso and banana variety Robusta. Materials and Methods Matured mangoes var. Alphonso were harvested by manual hand picking and latex was removed by keeping fruits in inverted position on the desaping bench. Then fruits were transported in plastic crates cushioned with newspapers to the laboratory. Similarly harvested banana var. Robusta fruits were hand separated from the main bunches. The fruits were then transported in plastic crates cushioned with newspapers to the laboratory. Sorting, Grading and Washing Fruits were sorted out for mechanical injuries like abrasions, punctures and bruises. Both mango and banana fruits were graded to maintain uniformity in the experiment. These fruits were washed in running tap water to remove the adhering latex, dust, and dirt, and excess moisture was allowed to drain off. The mango fruits were divided into five groups of 100 fruits each. Similarly banana fruits were divided into six groups of five hands each. Initial fruit field temperature was recorded with Tele-thermometer (YSI Model 43, yellow springs instruments USA) using 400 series temperature probes. Treatment Schedule Selected for the Experiment Mango Pre-cooling - By using forced air pre-cooler (Rinac India Ltd., Bangalore, India) T 1 - 8 ºC Pre-cooled air was used to pre-cool fruits to 8 ºC T 2 - 13 ºC Pre-cooled air was used to pre-cool fruits to 13 ºC. Hot water dip followed by cooling - By using hot water bath and forced air pre-cooler (Rinac India Ltd., Bangalore, India) T 3 - fruits were exposed to 52 - 55 ºC heat shock for 5 min followed by cooling to 8 ºC by using 8 ºC pre-cooled air T 4 - fruits were exposed to 52 - 55 ºC heat shock for 5 min followed by cooling to 13 ºC by using 13 ºC pre-cooled air T 5 - Control (Non-cooled and non-heated, fruit temperature was 28 ºC± 2 ºC). Banana Pre-cooling - By using forced air pre-cooler (Rinac India Ltd., Bangalore, India). T 1 - 5 ºC Pre-cooled air was used to pre-cool fruits to 8 ºC T 2 - 8 ºC Pre-cooled air was used to pre-cool fruits to 8 ºC T 3 - 13 ºC Pre-cooled air was used to pre-cool fruits to 13 ºC. Heat treatment followed by pre-cooling - By using hot water bath and forced air pre-cooler (Rinac India Ltd., Bangalore, and India) T 4 - fruits were exposed to 45 ºC heat treatment for one hour followed by cooling to 8 ºC by using 8 ºC pre-cooled air T 5 - fruits were exposed to 45 ºC heat treatment followed by cooling to 13 ºC by using 13 ºC pre-cooled air T 6 - Control (Non-cooled and non-heated, fruit temperature was 26 ºC± 2 ºC). Mangoes in the groups of 1, 2, 3, 4 and 5 were subjected to the treatments T 1 , T 2 , T 3 , T 4 and T 5, respectively. Similarly, fruits of banana grouped into six groups were subjected separately to the treatments T 1 , T 2 , T 3 , T 4 , T 5 and T 6, respectively. The temperature of the fruits was monitored by thermometer. The fruits were removed from the pre-cooler once the set temperature was reached. Fruits were observed for morphological injuries and immediately cut into small pieces followed by dipping in liquid nitrogen and storage at -40 ºC for further biochemical analysis. Extraction of Antioxidant Enzymes Five grams of frozen mango pulp and peel and pulp of banana were separately ground in liquid nitrogen using mortar and pestle previously chilled with liquid nitrogen and fine powder was used for extraction of antioxidant enzymes. Fine powder samples ground in liquid nitrogen were further extracted with 15 ml of ice cold sodium potassium phosphate buffer of pH 7.8 (0.1M) containing 5% (W/V) Polyvinylpyrrolidone (PVPP), 2mM dithiothreitol (DTT), 0.1mM ethylenediaminetetraacetic acid (EDTA) and 1.25 mM polyethylene glycol [14] . The homogenate was filtered through two layers of cloth and centrifuged at 20,000 X g for 15 minutes. The supernatant collected was used to determine the SOD and CAT activities. The extraction of total peroxidase (POX) was carried out using a previously published protocol [15] . The liquid nitrogen ground fine powder was extracted with 15 ml of ice cold sodium potassium phosphate buffer of pH 7.0 (0.1M) containing 0.5mM cysteine. The extract was centrifuged at 15,000 X g for 20 minutes. The supernatant collected was used for estimation of POX activity. Superoxide dismutase Assay The activity of SOD was assayed by measuring the ability to inhibit the photochemical reduction of nitroblue tetrazolium (NBT) using a published procedure [16] . Enzyme activity was measured spectrophotometrically at 560 nm. One unit of SOD was considered to be the amount of enzyme that inhibited NBT reduction by 50%. Catalase Assay The decomposition of H 2 O 2 was measured spectrophotometrically by measuring the change in absorption at 240 nm. CAT activity was assayed and calculated as one unit of CAT activity corresponded to the amount of enzyme that decomposes 1 µ mole of H 2 O 2 per minute [17] . Peroxidase Assay The POX activity was measured spectrophotometrically at 485 nm in reaction mixture consisting of 0.1 M potassium phosphate buffer pH 7.0, 12.5 mM o-phenylenediamine and 5 mM H 2 O 2 [18] . One unit of POX activity corresponds to the amount of enzyme responsible for change in absorbance of 1 absorbance unit minute -1 . All enzyme activities were expressed per g of fresh weight (FW). Data of enzyme activities measured was subjected to one-way ANOVA and means of five replicates were separated using the critical difference (CD) at 5% and depicted in the Figures. Results Different treatments of pre-cooling and heat resulted in no morphological injuries - chilling and heat injuries. Antioxidant enzymatic activity in Mango CAT activity Variation in the CAT activity after pre-cooling and hot water treatments followed by cooling of mango is presented in [Figure - 1]. Significant variations in the CAT activity in different treatments were observed. Heat-shock followed by cooling to 8 or 13 ºC induced the CAT activity. CAT activities in T 3 (12.52 µM H 2 O 2 min -1 g -1 FW) and T 4 (12.6 µM H 2 O 2 min -1 g -1 FW) treatments were significantly high (P≥0.05). Changes in CAT activity of the fruits of T 2 (5.70 µM H 2 O 2 min -1 g -1 FW ) and T 5 (5.98 µM H 2 O 2 min -1 g -1 FW) control treatments was non significant. Reduced CAT activity in response to cold stress in T 1 treatment was lowest (P≥0.05) as compared to other treatments [Figure - 1] (T 5 ). POX activity POX activity during pre-cooling and heat treatment followed by cooling to 8 ºC and 13 ºC is depicted in [Figure - 1]. POX activity in fruits kept under control conditions (T 5 ) was 0.51 units min -1 g -1 FW) and the changes in POX activity was found to be statistically non-significant in T 2 (0.5 units min -1 g -1 FW). However, increased POX activity in response to T 4 (0.65 units min -1 g -1 FW) and T 3 (0.64 units min -1 g -1 FW) treatments were significant. As it occurred with CAT, POX activity significantly (P≥0.05) declined in response to cold stress in T 1 (0.33 units min -1 g -1 FW) treatment, though it did not showed any chilling injuries. SOD activity Hot water treatment and followed by cooling to 13 ºC and 8ºC showed decreased SOD activity [Figure - 1]. Highest SOD activity of mango was measured in T 5 (119.4 units min -1 g -1 FW) treatment whereas least was recorded in T 4 (86 units min -1 g -1 FW). Treatments T 2 , T 1 and T 3 showed the SOD activity of 118.7 and 116.3 and 86.1 units min -1 g -1 FW, respectively. Antioxidant enzymatic activity in Banana The effect of pre-cooling and heat treatments followed by cooling to 8 or 13 ºC on the changes in ROS detoxifying enzymes CAT, POX and SOD of banana peel is shown in [Figure - 2]. As shown in [Figure - 2], control fruit (T 6 ) SOD activity was 2.84 units min -1 g -1 FW and reached to 2.94 units min -1 g -1 FW after pre-cooling to 13 ºC by using 13 ºC pre-cooled air (T 3 ) and it was non-significant (P≥0.05). Fruits subjected to treatments T 4 (0.4 units min -1 g -1 FW) and T 5 (1.8 units min -1 g -1 FW) recorded significantly (P≥0.05) decreased CAT activity and remained lower than T 1 (2.15 units min -1 g -1 FW and T 2 (2.02 units min -1 g -1 FW) treatments. Like banana peel CAT levels in T 3 and T 6 treatments, observed peel POX activity in T 3 (0.41 units min -1 g -1 FW) and T 6 (0.36 units min -1 g -1 FW) treatments were similar. The peel of fruits T 4 and T 5 has shown the POX activity of 2.1 and 1.91 units min -1 g -1 FW respectively. Heat stress followed by either cooling to 8 and 13 ºC in T 4 (2.1 units min -1 g -1 FW) and T 5 (1.91 units min -1 g -1 FW) induced the POX activity and found to be significantly higher than fruits subjected to cold stress alone T 1 (0.67 units min -1 g -1 FW) treatment. No variation in SOD activity in peel of banana subjected to different treatments when compared to untreated control fruit (T 6 ) [Figure - 2]. Peel of the T 2 and T 6 has shown the activity of 21.08 and 21.03 units min -1 g -1 FW respectively. The T 3 and T 1 showed 20.94 and 20.7 units min -1 g -1 FW of SOD activity, respectively, while it was 20.48 and 19.74 units min -1 g -1 FW, in T 5 and T 4 . Changes in the activities of ROS scavenging enzymes CAT, POX and SOD in banana pulp during pre-cooling and heat treatments followed by cooling to 8 or 13 ºC are presented in [Figure - 3]. CAT activity in the pulp of banana varied at different treatments than those in the peel [Figure - 2]. CAT activity of control fruit (T 6 ) was higher (16.72 µM H 2 O 2 min -1 g -1 FW) in general, than in other treatments. The activity of this enzyme declined significantly in response in the treatments T1 (5.44 µM H 2 O 2 min -1 g -1 FW) T 2 , (5.44 µM H 2 O 2 min -1 g -1 FW), T 4 , ( 8.76 µM H 2 O 2 min -1 g -1 FW) and T 5 . ( 13.04 µM H 2 O 2 min -1 g -1 FW) treatments and the decline was higher in T 1. ( 5.44 µM H 2 O 2 min -1 g -1 FW). However, a change in CAT activity was non-significant in T 3 (16.16 µM H 2 O 2 min -1 g -1 FW) at (P≥0.05). POX activity estimated in the pulp of the fruits subjected to different treatments had greater variation than in the peel [Figure - 3]. Highest activity was recorded in T 5 (8.02 units min -1 g -1 FW) followed by T 1 (7.91 units min -1 g -1 FW) and T 4 (7.17 units min -1 g -1 FW) respectively. Activity of 2.77 and 1.7 units min -1 g -1 FW was observed in the treatments T 2 and T 3 whereas in T 6 the activity recorded was 1.8 units min -1 g -1 FW. It is evident from the data that, the effect of T 3 (42.80 units min -1 g -1 FW) treatment on SOD levels compared with T 6 (43.20 units min -1 g -1 FW) of banana pulp was non-significant (P≥0.05). Increased SOD activity, during pre-cooling at 8 or 13 ºC and heat treatment at 45 ºC followed by cooling to 8 or 13 ºC was significant (P≥0.05). Highest SOD activity (49.60 unit min -1 g -1 FW) was observed when banana was subjected to treatment T 5 . Similarly, in the pulp of the fruit of T 4 resulted in the enzyme activity of 49.44 units min -1 g -1 FW followed by 49.20 units min -1 g -1 FW and 49.3 units min -1 g -1 FW, respectively in T 1 and T 2 treatment. Discussion Many plants and plant-products of tropical origin suffer injury and even death when exposed to low but non-freezing temperatures. Chilling sensitive fruits and vegetables will develop various characteristic symptoms, or disorders, when exposed for sufficient periods of time at temperatures below 15 °C, depending on the species, cultivar and tissue type. These disorders include: pitting, abnormal ripping, necrosis and discoloration and increased disease susceptibility [19] . During visual assessment of mango and banana fruits after pre-cooling (using different temperatures) no chilling injury (CI) symptoms were observed even though the storage temperatures were 5 °C and 8 °C. Storage treatments influenced antioxidant enzymes activities and variations were recorded which could be due to very short exposure period. Similarly, previous reports recommended 13 °C as suitable for storage of banana and mango [20] . Post harvest dips to control decay have been often practiced only for few minutes and treatments at higher temperatures in hot water dip / heat treatments have been practiced to kill insect pests located at the interior of the commodity because only the surface of the commodity requires heating. In the present study, application of 52-55 °C (generally adopted as quarantine measures) for 5 minutes to mango has not shown any fruit damage due to heat injuries like skin scalding, damaged lenticels which could be due to very short exposure period and cultivar specific unlike the results recorded while working with post harvest handling of fruits and vegetables [21] . Generally, plants show a rapid response to increased AOS produced by abiotic stresses like changes in storage temperature [22] . These changes lead to an enhancement in cellular antioxidants that induce an increase in the activity of natural antioxidants and antioxidant enzymes such as SOD, CAT, GR and APX [23] . In the present investigation, CAT activity measured after T 3 and T 4 (52 ºC - 55 ºC heat treatment for 5 min followed by cooling to 8 ºC and 52 ºC - 55 ºC heat treatment for 5 min followed by cooling to 13 ºC) treatments to mango were significantly higher than those under T 1 , T 2 and T 5 (pre-cooling at 8 ºC, 13 ºC and control) treatments. Peroxidase activity in the pulp of the mango treated with T 3 , T 4 and T 5 (heat shock followed by cooling at 8 ºC and 13 ºC and control) were significantly different from those of T 1 and T 2 (pre-cooling at 8 ºC and 13 ºC). The results found in the present investigation show that applied heat treatment, followed by cooling to low temperature, may increase the capacity of Alphonso mango fruit to remove an excess of hydrogen peroxide by CAT and POX. Our results indicate that heat treatment followed by cooling did not enhance the ability of Alphonso mango to dismutate superoxide radicals, as the activity of the enzyme SOD was lower in the heat-treated fruit. This may be due to heat-induced ethylene in association with low temperature that would have lowered the SOD activity. These results are in corroboration with the observations made indicating that ethylene, in association with low temperature, lowered the SOD activity [24] . Whereas SOD activity differences in the T 1 , T 2 and T 5 (pre-cooling at 8 ºC and 13 ºC and control) were significantly different from those of T 3 and T 4 (heat shock followed by cooling at 8 ºC and 13 ºC). Antioxidant enzyme CAT activity in the pulp was higher than that in the peel of the banana fruits subjected to the pre-cooling and heat treatments. Activity measured in the pulp and peel of the fruits treated with T 3 (13 ºC PC to 13 ºC) and control was significantly different from that of other treatments T 1 , T 2 , T 4 and T5 (pre-cooling at 5 ºC, 8 ºC, heat treatment followed by cooling at 8 ºC and 13 ºC). Unlike CAT activity, peroxidase activities in the peel and pulp were varying independently. Enzyme activity in the peel of the fruits subjected to the treatment of heat followed by cooling to 8 ºC was higher and significantly different from that of other treatments whereas the activity in the pulp of the fruits treated with pre-cooled at 5 ºC to cooling at 8 ºC and quarantine heat treatment followed by cooling at 13 ºC was significantly higher than those at other treatments. POX plays also role in the breakdown of hydrogen peroxide, and therefore post harvest chilling and heat induced increase in POX that could be related to the defense mechanism of fruit to cope with the oxidative stress. The affinities of CAT (µM range) and other Peroxidases like Ascorbate peroxidase (mM range) for hydrogen peroxide are very different which belong to two classes of hydrogen peroxide-scavenging enzymes. Thus, although CAT is involved in the removal of excess reactive oxygen species (ROS) during stress conditions in plants, the other peroxidases like Ascorbate peroxidase, which has been reported to be one of the most important antioxidant enzymes in defense against low-temperature injury, appears to be responsible for the modulation of ROS for signaling [25] . The results showed that the activity of total POX increased during pre-cooling at low temperatures (8 and 13 ºC) and heat treatment followed by cooling to 8 or 13 ºC. Statistically, SOD activity measured in the peel of the fruits treated with different treatments was not significant. Superoxide dismutase activity in the pulp of the fruits treated with pre-cooling at 5 ºC, 8 ºC, heat treatment followed by cooling to 8 ºC and 13 ºC was significantly higher and was different from that in those fruits subjected to other treatments and control. Thus results showed heat related oxidative stress, pre-cooling at 5 or 8 ºC and heat treatments followed by cooling to 8 or 13 ºC up regulated the activity of SOD. This can be an important protection mechanism of pulp of Robusta banana against the excessive increase of ROS during post harvest pre-cooling and heat treatments. Conclusion These results highlight the Alphonso mango and Robusta banana capacity to withstand various post harvest pre-cooling and heat stress conditions. Active oxygen species were generally produced by enzymatic and non-enzymatic pathway during cellular metabolism. The present study showed that the level of expression of antioxidant enzymes during pre-cooling and heat treatment conditions dependent upon exposed temperature and underline the important role for protecting cellular apparatus during pre-cooling and heat treatments. Extent of recommendation of pre-cooling temperature and heat treatment of these fruits after pre-cooling at 5 ºC and 8 ºC can be made based on the levels of reactive oxygen species. Research into variations in the activities of other antioxidant enzymes and the action of other molecules with a non-enzymatic action may give a more complete picture of the response of Alphonso mango and Robusta banana fruits to postharvest oxidative stress and better explain its high resistance to this specific abiotic stress. Therefore, findings of the present study serve as basic information about cellular homeostasis in the mango and banana during pre-cooling and heat treatments. Acknowledgement The authors wish to thank the Director, IIHR Bangalore for providing facilities, encouragement and support. Authors are also thankful to Dr(s). S K Jalali, P Sharanappa and Y Lalitha for inputs in carrying out statistical analyses and their useful comments. References
Copyright 2009 - African Journal of Food Agriculture, Nutrition and Development The following images related to this document are available:Photo images[nd09056f3.jpg] [nd09056f2.jpg] [nd09056f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}