 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
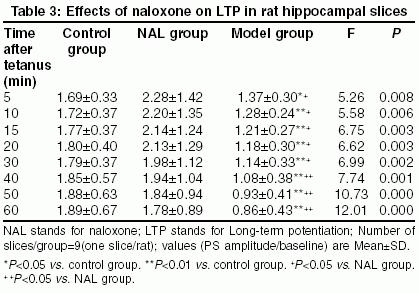
Neurology India, Vol. 53, No. 1, January-March, 2005, pp. 79-82 Original Article Effect of naloxone on aluminum-induced learning and memory impairment in rats Shi-lei Sun, Guang-yu MA, Bachelor LIHua, Bachelor ZhuYun, Hong-mei Dong, Xiao-hu XU Department of Gerontology, First Affiliated Hospital, Zhengzhou University, ZhengZhou - 450 052 Code Number: ni05023 Abstract BACKGROUND: Uptake of aluminum may disturb the learning and memory of humans or animals. Naloxone (NAL) has been shown to exert beneficial effects on memory deficits.AIMS: We investigated the effects of naloxone on aluminum-induced learning and memory impairment in rats. SETTINGS AND DESIGN: Aluminum-induced learning and memory impairment model was established by gavage of Aluminum chloride (600 mg/kg) for 3 months. Rats were divided into three groups viz. naloxone-treated rats (NAL 0.8 mg/kg, i.p. daily for 7 days), non-treated model rats and normal controls. MATERIALS AND METHODS: The Morris water maze test was performed to study spatial learning and memory. Long-term potentiation (LTP) of the Schaffer collateral-CA1 synapse was recorded. Aluminum and zinc contents in the hippocampus were assayed with atomic absorption spectrophotometry. STATISTICAL ANALYSIS: Parameters of the hidden and visible platform trials and data of LTP were analyzed using two-way repeated measures ANOVA. RESULTS: In the hidden platform trials, escape latencies of the NAL rats were significantly shorter than that of the non-treated rats (P=0.000, 95% confidential interval low bound 14.31, upper bound 22.68). In probe trails, the number of entries in the target area of the NAL rats (6.75±1.28 times/min) was more than that of non-treated model rats (4.56±2.16 times/min, P=0.004, 95% confidence interval low bound -3.65, upper bound -0.788). The magnitudes of LTP recorded in the CA1 pyramidal neurons of the NAL-treated rats were significantly augmented when compared to the non-treated model rats (P=0.005, 95% confidence interval low bound 0.16, upper bound 0.84). CONCLUSIONS: NAL could facilitate spatial learning and memory and enhance LTP in the CA1 region of the hippocampus in aluminum-induced learning and memory impairment in rats. Key Words: Naloxone, Aluminum, hippocampus, learning and memory, long-term pontentiation, Morris water maze Introduction More and more evidence has implicated aluminum (Al) as a neurotoxin,
which is involved in the etiology of many neurodegenerative disorders,
such as Alzheimer′s disease (AD) and dialysis dementia.[1] AD manifests with insidious and progressive learning and memory loss. It has been reported that the endogenous opioid system in the central nervous system could significantly impair learning and memory, which can be reversed by naloxone (NAL), an opioid receptor antagonist.[2] Materials and Methods Animal treatments: All animal treatments were conducted in accordance with the European Communities Council Directive of 24 November 1986 (86/609/EEC). Before the experiment, 27 male and female Sprague-Dawley rats (body weight, 379.1±20.5 g, certification No. 24101101 conferred by Medical Animal Management Committee, Sichuan Province) were housed for one week for habituation, three animals per cage in a temperature-controlled environment (20-24 oC) under a 12:12 h light/dark cycle.Rats were gavaged tap water containing Aluminum chloride (600 mg/kg), into the stomachs daily for 3 months. The containers or tubes were washed with acid. Then rats were randomly divided into two groups viz. NAL-treated model rats and non-treated model rats. Nine rats formed the normal control group. Rats in the NAL group received i.p. injection of NAL (0.8 mg/kg), daily for 7 days (Days 1-7), and the other group received injection of an equal volume of normal saline solution at the same time. From Day 8 the Morris water maze training was carried out. Morris water maze training The Morris water maze comprised a circular water bath (2.0 m in diameter) filled with tap water to a depth of 0.3 m (22-23oC) and made opaque. A Perspex platform (11 cm in diameter) could be placed in any quadrant of the bath. Each trial started by placing the rat gently in the water, facing the edge of the water bath, at the designed location.[5],[6] The Morris water maze training comprised four components. a. Pre-training trials (Days 8-10) - To familiarize the rats with the maze and the escape procedure. b. Hidden platform trials (Days 11-16) -Each rat was given four consecutive trials per day for 6 days. Starting positions from arbitrarily assigned compass locations were randomized for each rat on each day. The platform was kept in a constant position for all rats. The time taken to reach the platform (escape latency) was noted. c. Visible platform trials (Days 17-18) - Rats were given four trials per day (inter-trial interval 3-8 min) for 2 days with the platform (protruded 1 cm above the surface of the water) in each of the four quadrants with the same start location and escape latency recorded in each trial. d. Probe trials - On Day 16, 4 h after the last hidden platform trial, the platform was removed and the rats were allowed to swim for 1 min and the number of entries in the target area where the platform used to be was noted.[7] Slice preparations: On Days 19-22, 12 h after the last water maze training, rats were decapitated one by one at intervals of at least 100 min. The brain was quickly removed. The hippocampus was dissected out and cut into transverse slices 400-µ m thick. The fresh slices were incubated for at least 90 min at 33oC in artificial cerebrospinal fluid (aCSF pH 7.4) of the following composition (in mM): NaCl, 124; KCl, 3.3; KH2PO4, 1.2; MgSO4. 7H2O, 1.5; CaCl2, 2.4; NaHCO3, 25; D-glucose, 10; bubbled with warm gas (95% O2+5% CO2). Electrophysiology: For extracellular recording, a glass electrode was inserted in the pyramidal layer of the CAl region for the recording of field potential (population spike, PS). A bipolar stimulating electrode was placed in the Schaffer collateral-commissural fiber to activate the LTP. Before the production of tetanus, the population spike evoked by a single pulse was recorded for 15 min as baseline. Rectangular pulses were delivered at 100 Hz through the stimulating electrode, as test stimulation, at a strength that gave 70 per cent of the maximal response.[5] Tetanization parameters: LTP was induced by a theta burst stimulation consisting of 3 bursts of 50 pulses (100 Hz frequency, 0.2 msec stimulus duration, 10 msec inter-burst interval). The stimulus intensity was the same as that used for the recordings. Data analysis: The amplitude of the PS was measured as the distance from the negative peak to the positive. The magnitude of LTP in each preparation was expressed as 20 per cent increase of amplitude of the averaged responses. The data were expressed as a ratio of PS amplitude to baseline. Determination of Al and Zn: The Al and Zn content in the hippocampus was determined by atomic absorption spectrophotometry.[8] Statistical analysis: All data were expressed as mean±SD. For the hidden and visible platform trials the latencies of each rat on each day were averaged prior to analysis.[6] These parameters and data of LTP were then analyzed using two-way repeated measures ANOVA (RM-ANOVA) with SPSS 10.0 with treatment (NAL) as the inter-subject variable and the time (days or time point after tetanus) as the intra-subject variable. Post hoc comparisons were made using Dunnett′s test (all groups compared to control) and Tukey′s test (all groups compared with each other). For the probe trial and the Al, Zn content, the data were analyzed using a one-way ANOVA with multiple comparisons as described above. Results Al and Zn content in the hippocampus: The Al content of non-treated model rats (13.22±1.47 μg/kg) and NAL-treated rats (10.90±1.39 μg/kg) was elevated as compared to that of normal controls (4.53±0.23 μg/kg) (F2,24=58.53, P=0.000), while there was no significant difference between the former two. There was no significant difference in the Zn content among the three groups (F2,24=0.91, P=0.43).Hidden platform trials (Days 11-16): Two-way RM-ANOVA showed a significant effect of NAL treatment on the reduction of escape latency (F2,24=62.303, P=0.000), [Table - 1]. Generally, escape latency was significantly longer in the non-treated model rats compared to NAL rats (P=0.000, 95% confidence interval low bound 14.31, upper bound 22.68, two-way RM-ANOVA followed by Tukey′s test) and controls (P=0.000, 95% confidence interval low bound 17.65, upper bound 26.01, two-way RM-ANOVA followed by Dunnett′s test). On Day 5 and Day 6 trials the escape latency of NAL rats was longer than that of normal controls, while in other trials no significant difference was found (P=0.636, 95% confidence interval low bound -0.85, upper bound 7.52, two way RM-ANOVA followed by Dunnett′s test). Visible platform trials (Day 17-18): Two-way RM-ANOVA demonstrated no significant effect of NAL on escape latency in rats in the visible platform trials (F2,24=1.300, P=0.291, [Table - 2]. Probe trials: One-way ANOVA showed a significant effect of NAL treatment on the number of entries in the target area in rats (F2,24=7.991, P =0.002). The number of entries was significantly increased in the NAL-treated rats (6.75±1.28 times/min) compared to the non-treated rats (4.56±2.16 times/min, P=0.004, 95% confidence interval low bound -3.65, upper bound -0.788, Tukey′s test). There wasn′t a significant difference in the number of entries between the NAL-treated rats and the normal controls (7.81±2.12 times/min, P=0.636, (P=0.000, 95% confidence interval low bound -1.77, upper bound 1.10, Dunnett′s test). LTP in hippocampal slices of rats: LTP was studied in rats that had been tested in the water maze (n=9 slices/group, one slice/rat). After tetanic stimulation, the PS amplitudes increasing more than 20 per cent compared to the mean amplitude before tetanus and lasting more than 30 min suggested that LTP have been induced in the CA1 region. Also the increased PS evoked by high frequency stimulation in the controls and the NAL-treated rats was seen to be significantly elevated as compared with that of the non-treated model rats (P=0.005, 95% confidence interval low bound 0.16, upper bound 0.84, Tukey′s test). No significant difference in PS amplitude was found between the NAL rats and the normal controls (P=0.328, 95% confidence interval low bound -0.43, upper bound 0.15, two way RM-ANOVA followed by Dunnett′s test). Two-way RM-ANOVA showed a significant effect of NAL treatment on PS amplitude (F2,24=7.408, P=0.002) [Table - 3]. Discussion Uptake of Al, from Al-treated drinking water over a long period time, may directly enter the brain tissue and disturb the memory and learning of humans or animals.[9] In the present hidden platform trials, the latencies of the non-treated model rats were significantly longer than that of the normal controls, the number of entries in the target area in probe trials was significantly decreased, and the magnitudes of LTP were significantly decreased, which showed that long-term (3 months) intake of Al could induce learning and memory deficits in rats. Acknowledgment This work was supported by the Natural Science Foundation of Guangdong Province, ChinaReferences
Copyright 2005 - Neurology India The following images related to this document are available:Photo images[ni05023t1.jpg] [ni05023t3.jpg] [ni05023t2.jpg] |
| |||||||||

![[Table - 1]](/showimage?ni/photo/ni05023t1.jpg){kind=link}
![[Table - 2]](/showimage?ni/photo/ni05023t2.jpg){kind=link}
{kind=link}