 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
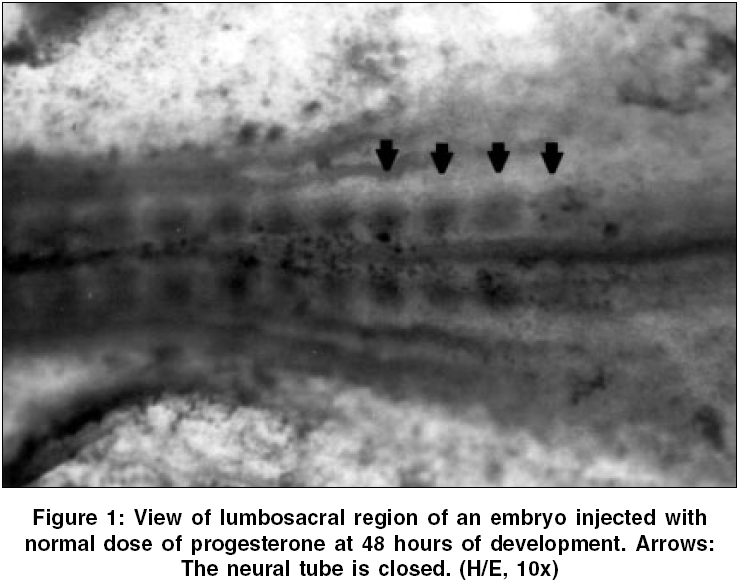
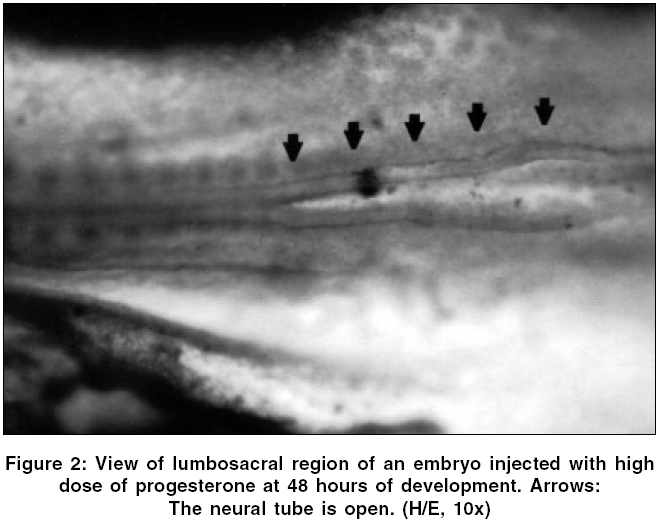
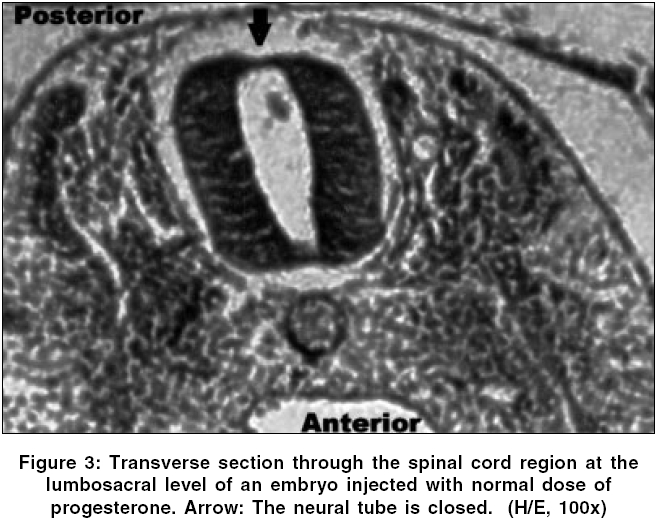
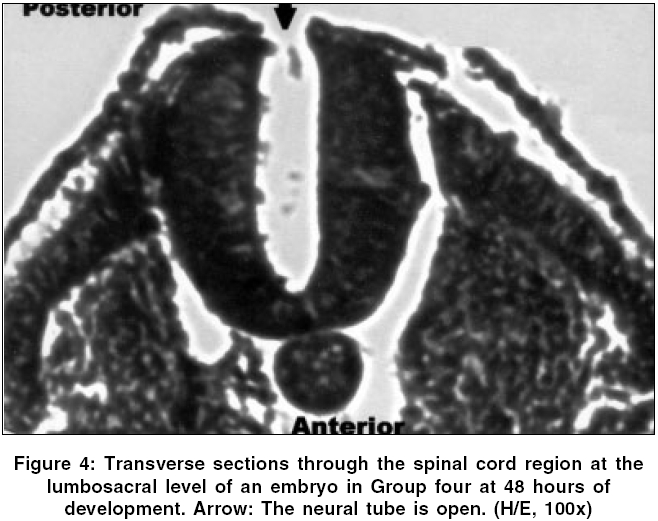
Neurology India, Vol. 54, No. 2, April-June, 2006, pp. 178-181 Original Article The effects of high dose progesterone on neural tube development in early chick embryos Pamir Erdincler, Ali Dalgic, Ismail Cepni, Sanli Erkan, Ali Kafadar, Mustafa Tasyurekli Departments of Gynecology and Obstetrics, Istanbul University, Cerrahpasa Medical Faculty, Istanbul Code Number: ni06052 Abstract Background: Although folic acid deficiency is known to be one of the factors in the development of spina bifida and other neural tube defects (NTD) the exact pathophysiology still remains unclear. Progesterone is an endogenous hormone which increases significantly during pregnancy.Aims: We aimed to study the possible negative effects of high dose progesterone on neural tube development in early chick embryos. In order to test our hypothesis, early chick embryos were exposed to physiological saline, normal and high doses of progesterone. Settings and Design: 160 fertile, specific pathogen free white leghorn eggs (Gallus gallus), all at stage eight of development were divided into four equal groups. Materials and Methods: The first group was incubated without any operation. The second group was injected with physiological saline. The third and fourth groups were injected with two and twenty times more than physiologic doses of progesterone respectively. After 48 hours of incubation, all embryos were analyzed for the presence of NTDs under light microscopy. Statistical Analysis Used: None. Results: At 48 hours of incubation, 84% (135/160) of the embryos passed characteristics of Stage 12 development and were included to the study. None of the eggs in the first three groups showed NTDs, whereas 81.8% (27/33) of the eggs in the fourth group showed NTDs. Conclusions: Our study showed that progesterone at levels twenty times more than its physiologic level might cause NTDs. Further studies are needed to explain the mechanisms of this teratogenic effect. Keywords: Neural tube defects, progesterone, chick embryo, neural tube closure, spina bifida. Introduction One of the early and important events in the development of central nervous system is the closure of the neural tube. Defects in this closure process lead to a number of malformations, which can create problems in the developing fetus.[1]Progesterone is a 21-carbon (C 21 H 30 O 2 ) steroid hormone that is associated with pregnancy. It is also widely used in "in vitro fertilization" therapies.[2] The aim of this study is to determine whether an endogenous molecule such as progesterone could have adverse effects on the development of the neural tube in early chick embryo models with the help of histopathologic changes under light microscopy. Materials and Methods Chick embryos Fertile, specific pathogen free eggs of the domestic fowl (White Leghorn, Gallus gallus ) were obtained from Manisa Celal Bayar University, Research Institute of Poultry Disease and Vaccination, at zero hour of incubation. The eggs were incubated at 37.5° C and 75% relative humidity until the embryos reached stage eight of development according to Hamburger and Hamilton.[3] At this stage the eggs were divided into four groups consisting of 40 eggs per group: group one, uninjected eggs; group two, sham-operated eggs (injected with physiological saline); group three, injected with twice the normal dose of progesterone; and group four, injected with high dose (twenty times more than physiologic level) of progesterone. Dosage of progesterone The normal progesterone level that a chick embryo is exposed to is found to be 0.823±0.035 nanogram/ml.[4] Twice the equivalent dose of this level of progesterone (water soluble progesterone: (C 21 H 30 O 2 ) Sigma - Aldrich Comp. code: P7556, St.Louis, Missouri, USA) per egg was calculated to be 15.7 nanograms. The calculated dose of progesterone was diluted in 0.1 ml of physiological saline (0.9% NaCl) for group three. For high dose of progesterone group (group four), 157 nanograms of progesterone was injected in the same volume of physiological saline. Method of injection At stage eight of development, the eggs were washed with 70% alcohol and properly labeled on the outer shell. A hole was made on the blunt pole of the eggs with a sharp and thick needle under laminar flow. Using a sterile 28-gauge needle and a tuberculin syringe, 0.1 ml of the fluid was injected from the blunt end under the embryonic disc. The holes were sealed with paraffin. The eggs were then placed in an incubator. Embryo collection The opening of the eggs was done at 48 hours of incubation. The eggs were cracked open and the outer shell was chipped out to create a large opening to see the embryo. The viability of the embryos was assessed by the heart beat. The embryos were transferred to a petri dish by careful dissection among the allantoic stalk and other embryonic structures. All the embryos were fixed with 10% formalin and stained with hematoxylin and examined under stereomicroscope to assess any gross developmental abnormalities.[5] Then, embryos which passed Hamburger Hamilton stage 12 (34, 35, 33 and 33 in groups 1, 2, 3 and 4 respectively) were embedded into paraffin and five microns thick paraffin sections were cut and stained with hematoxylin and eosin for light microscopic examination.[5] Results At the time of injection, each embryo had four somite pairs and the neural folds of the future midbrain and a portion of the hindbrain had already made contact. After a further 22 hours of post injection incubation, 25/160 (15.5%) of the embryos showed no or insufficient development. 34 / 40 (85%) embryos of the group one (uninjected), 35 / 40 (87.5%) embryos of the group two (injected with physiological saline 0.9% NaCl), 33 / 40 (82.5%) embryos of the group three (injected with twice the normal dose of progesterone) and 33 / 40 (82.5%) embryos of the group four (injected with high dose of progesterone) passed characteristics of Stage 12 development. Several features that define this stage are: head turning to left side; anterior neuropore closure, telencephalon identifiable; primary optic vesicles and optic stalk well established; auditory pit deep but wide open; heart slightly S-shaped; and head-fold amnion covering the entire forebrain region.[3] In groups one, two and three; the neural tube was closed in all of the embryos which passed Stage 12. [Figure - 1] However, 27 / 33 (81.8%) embryos in group four which passed Stage 12 were malformed. [Table - 1] In these malformed embryos, mesenchyme and development of the organ systems was appropriate for their stage and the rostral part of the neural tube did not differ significantly from other normal embryos. However, the caudal part of the neural tube, corresponding to the lumbosacral regions of the embryos, was defective. [Figure - 2] In groups one, two and three, transverse sections through the spinal cords at the lumbosacral level showed neural folds in contact with each other, [Figure - 3] whereas the embryos in group four showed widely open neural folds. The defects were such that the neural folds showed no signs of contact throughout the neuroepithelium. [Figure - 4] Discussion Most of the neural tube defect (NTD) cases are thought to be a result of a combination of genetic predisposition and environmental trigger factors. Although few of the environmental causes and none of the genetic factors have been identified in humans so far, numerous teratogens and nutritional deficiencies have been suggested as possible causative factors. Recently, it was reported that periconceptional vitamin supplementation was associated with a statistically significant reduction in NTD cases when compared with an unsupplemented group.[6]Studies on the teratogenic effects of progesterone have dealt with possible cardiac malformations, the effects on the central nervous system, development of amelia, but mostly with the possible masculanizing effect on the female fetus.[7],[8] The data emerging from these studies have often been contradictory and inconclusive, mainly because of the lack of uniformity of the study material and the methodology. The early chick embryo model is an ideal model that corresponds to the first month of embryonic development in mammals and is well suited to the investigation of the effects of chemicals on the development of embryos. Numerous chemical agents such as caffeine, phenytoin, diazepam and local anesthetics are known to cause neural tube defects in chick embryos.[9] Stage eight embryos were generally chosen in these studies for the investigations because the developing neural tissue exhibits a gradual variation in the degree of opening along its length which provides an excellent opportunity to study the effects of chemical agents on closure of the neural tube.[10] The teratogenic effect of the large dose of progesterone might also be a chemical effect unrelated to its biological function. In either case, results obtained from our study on chick embryos indicate that high dose of progesterone exerted an inhibitory effect on the closure of neural tube. The normal function of progesterone in reproduction in birds is to induce the synthesis of egg-white proteins, including ovalbumin, conalbumin, lysozyme and ovomucoid by binding to the genes encoding these proteins.[11] Because of its water soluble properties, it is one of the few metabolites, which can pass through the protective barrier of the embryo.[12] In other words, the embryo, in a stage in which it is very sensitive to adverse effects of environmental factors, has a close relation with progesterone. How progesterone causes NTDs was not studied in this study, but some speculative explanations are available in the literature. It is known that progesterone acts in the brain by stimulating GABA signaling pathways in specific areas of the brain.[11] There is also evidence to suggest that substances which alter the function of the GABAergic system may contribute to the formation of neural tube defects.[13] Given that GABA is important to neural development and the early developmental time frame of the GABA system, it is logical that agents active at the GABA receptor such as progesterone can have adverse consequences on central nervous system development. A recent study by Brent concluded that progestational agents did not result in nongenital malformations.[14] Although this seems to refute the interpretations in this manuscript, our results showed that progesterone might cause neural tube defects in twenty times more than its physiologic level. In conclusion, the findings of this study suggest that administration of very high amounts of exogenous progesterone during early pregnancy could have deleterious effects on neural tube closure. Replication of this experiment and further studies are also needed to explain the mechanisms of this teratogenic effect. References
Copyright 2006 - Neurology India The following images related to this document are available:Photo images[ni06052f3.jpg] [ni06052f4.jpg] [ni06052f2.jpg] [ni06052t1.jpg] [ni06052f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}