 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
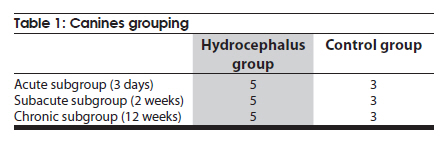
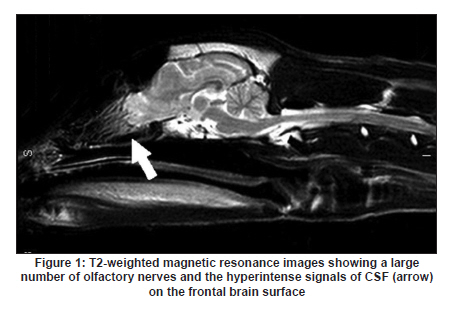
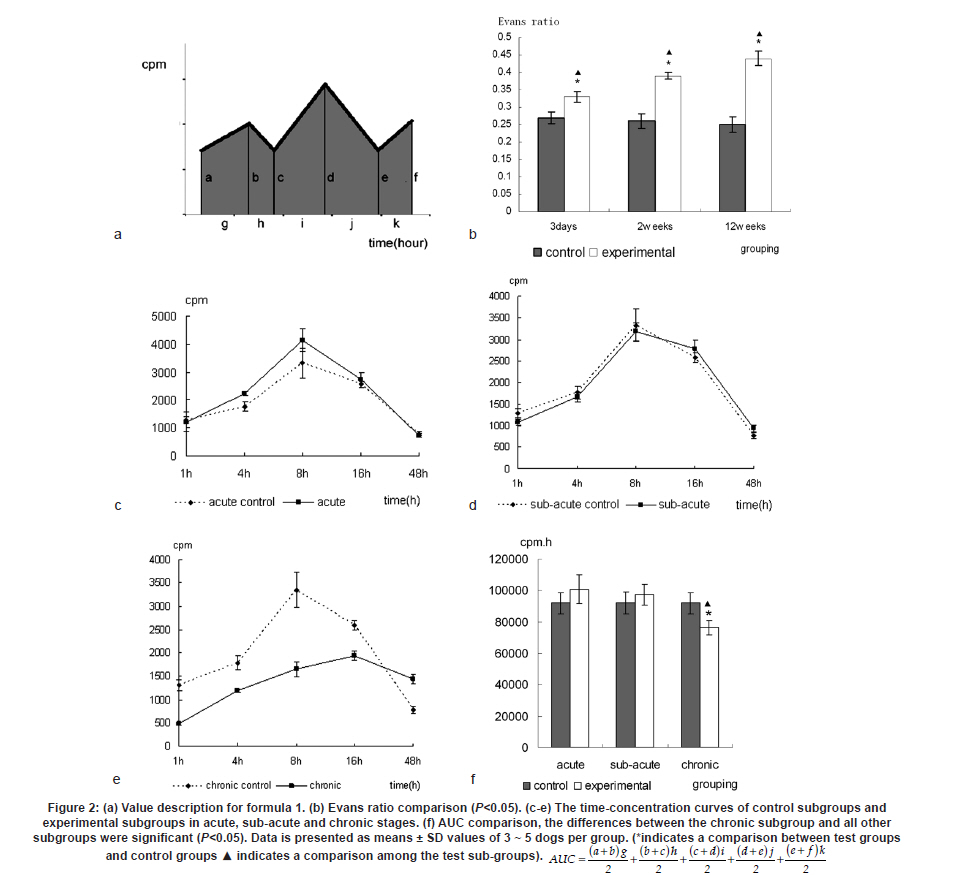
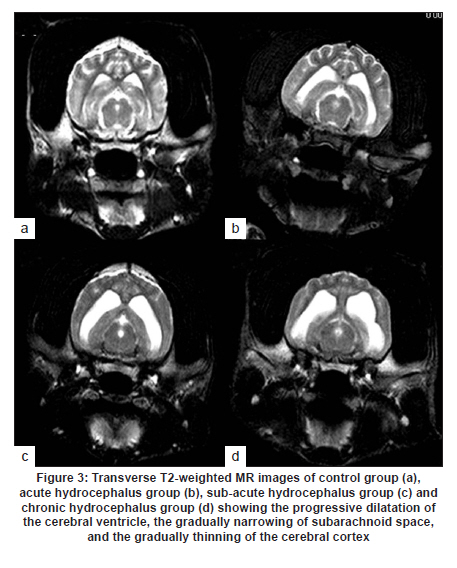
Neurology India, Vol. 58, No. 3, May-June, 2010, pp. 371-376 Original Article Cerebrospinal fluid absorption disorder of arachnoid villi in a canine model of hydrocephalus Ke Zhao1, Hong Sun1, Yan Shan2, Bo Yong Mao1, Heng Zhang1 1 Department of Neurosurgery, West China Hospital, West China School of Medicine, Sichuan University, Chengdu, Sichuan - 610 041, P. R, China Date of Acceptance: 10-Jun-2010 Code Number: ni10099 PMID: 20644263 Abstract Background: Hydrocephalus results from inadequate passage of cerebrospinal fluid (CSF) from its point of production within the cerebral ventricles to its point of absorption into systemic circulation.Aims: The objective of this study was to investigate the disorders of CSF absorption by arachnoid villi during the different phases of hydrocephalus. Materials and Methods: Silicone oil was injected into the fourth ventricle of 15 canines as an experimental group. Saline solution (0.9% NaCl) was injected in another nine canines as a control group. In order to block CSF transport through the cribriform plate, an external ethmoidectomy was performed in five dogs from experimental group and three dogs from control group at three days (acute stage), two weeks (sub-acute stage), and 12 weeks (chronic stage) respectively. Tritiated water was injected into the canines' cortical subarachnoid space and blood levels were measured at intervals of 1h, 4h, 8h, 16h and 48h respectively. Time-concentration curve of tritiated water was drafted. The area under the curve (AUC) was calculated for variance analysis and t-testing. Results: In the chronic group, the tritiated water concentration rose slowly to a peak at 16h. It was significantly lower than other groups at 1h, 4h, 8h and 16h, but was higher than other groups at 48h. Analysis of the AUC showed significant differences among all the groups (P<0.01). There were no significant differences in the AUC between control groups, the acute group, and the sub-acute group (P>0.05); however, the AUC of the chronic group was significantly lower than other groups (P<0.05). Conclusions: The CSF absorption ability of arachnoid villi is significantly damaged in a long-term state of hydrocephalus. Keywords: Arachnoid villus, cerebrospinal fluid, hydrocephalus, tritiated water Introduction Hydrocephalus is an active distension of the ventricular system of the brain resulting from inadequate passage of cerebrospinal fluid (CSF) from its point of production within the cerebral ventricles to its point of absorption into systemic circulation. [1] Hydrocephalus is a common neurosurgical disease often resulting from intracranial hemorrhage, tumors, intracranial infection, brain injury, and craniotomy. [2],[3] Abnormalities of CSF secretion, circulation, and absorption can lead to excessive accumulation of CSF in the ventricular system and the development of hydrocephalus. [4],[5],[6] Disturbances of CSF absorption play an important role in the development of hydrocephalus. [7],[8] Arachnoid villi are the predominant point of CSF absorption. [9],[10] Canine's arachnoid villi are mainly distributed around the superior sagittal sinus. This structure is similar to that of human arachnoid villi. [11] In a previous study we induced a adult mongrel canine model of hydrocephalus by silicone oil injected into the fourth ventricle. Electron microscopic and immunofluorescent staining studies showed that chronic hydrocephalus (at 12 weeks of experiment) could lead to subsequent morphological changes of the arachnoid villi. [11] From our previous results we hypothesized that the morphological changes of the arachnoid villi may result in a dysfunction of CSF absorption. In this follow up study, CSF absorption by the arachnoid villi was investigated in the different stages of hydrocephalus in canine models with the aid of isotope tracing after blocking the main lymphatic drainage pathway of CSF. Materials and Methods Animals Twenty-four healthy adult mongrel canines (1-2 years old, 10-15 kg) without ventriculomegaly, purchased from Huaxi Laboratory Animal Center, Sichuan University, China, were used in this study. Dogs were fed mixed feed and water ad libitum. Experiments were approved by ethics committee and the Institutional Committee for Animal Care, Sichuan University, China. See [Table - 1] for a detailed list of randomized animal groups. Induction of hydrocephalus model The dogs were fasted for 12h before surgery. All experimental canines were fixed in a sitting position on the operating table following anesthesia with intravenous pentobarbital (30 mg/kg). Straight incisions (10 cm) were made from the external occipital protuberance to the spinous process of the third cervical vertebrae. The foramen magnum regions were surgically exposed; a puncture hole (about 2 mm) was made at the midline of posterior atlantooccipital membrane. A silicone catheter (diameter 1 mm) was inserted with its tip directed toward the fourth ventricle. After slowly advancing the catheters about 15 mm without resistance, the catheters entered the fourth ventricles. CSF was drained via the silicone catheters (0.3 ml/kg) and was simultaneously replaced by equivalent amounts of Silicone oil (Dow Corning 200® Fluid; viscosity: 10,000 Centi Stokes (Cs)) or Saline solution (0.9% NaCl) by slow injection (1ml/min) in experimental and control groups respectively. After removing the silicone catheters, the puncture holes were covered with gelfoam and the incisions were sutured tightly. Evaluation of hydrocephalus Magnetic resonance imaging (MRI) of the head and brain was acquired in both groups of canines (control and test) at 3 days, 2 weeks, and 12 weeks. The dogs were anesthetized with intravenous pentobarbital (30 mg/kg). Baseline transverse and sagittal MR images (750/20 [TR/TE]; section thickness: 2.0 mm; intersection gap:1.0 mm) were acquired by a 3.0 Tesla MRI Scanner. We measured the widest internal transverse diameters of the skulls (AB) and the widest transverse diameters of the anterior horns of the lateral ventricles (CD) from T2-weighted images. Evans ratio (CD/AB) was used for assessment of the degree of hydrocephalus. Blocking CSF absorption through the cribriform plate In order to block CSF transport through the cribriform plate [Figure - 1], an external ethmoidectomy and sealing was performed on all the dogs according to their predetermined hydrocephalus model subgroups at 3 days, 2 weeks, and 12 weeks respectively. The skin over the frontal-nasal area was reflected to reveal the frontal and nasal bones. A portion of the nasal bone approximately 3Χ3 cm 2 was removed to expose the nasal mucosa. To effectively block nasal mucosa lymphatic drainage, the nasal mucosa, olfactory nerves, and all soft tissue on the extracranial surface of the cribriform plate was scraped away with a curette, and the bone surface was sealed with bone wax. Tritiated water injection and determination A burr hole was made approximately 10 mm caudally and 8mm laterally to the external occipital protuberance. Following the dissection of the cerebral dura mater, silicone catheters (diameter 1mm) were carefully inserted. One milliliter of tritiated water (including 3 H 2O 50μCi, Institute of Isotopes, China Institute of Atomic Energy) was injected into the canine cortical subarachnoid space when CSF flowed out. After injection of tritiated water, venous blood (2 ml) was sampled from jugular vein, respectively, at 1h, 4h, 8h, 16h and 48h. It was necessary to allow the sample tubes to stand vertically for the serum separation afterwards. The tritiated water content in the samples was measured using liquid scintillation counts by a FJ-2107P automatic change liquid scintillation counter (Xian Nuclear Instrument Factory, China). [12] Data processing and statistical analysis All data processing was done using Statistical Package for Social Sciences (Windows v11.5, SPSS Inc., Chicago, IL). All values were expressed as mean. The tritiated water time-concentration curve was drafted to describe the absorptive characteristics in each group according to the data. We calculated the area under the curve (AUC) using the Trapezoidal method (Formula 1, [Figure - 2] Value Description), then the results were analyzed by analysis of variance (ANOVA) and t-tests. P<0.05 was considered statistically significant. Results Model of obstructive hydrocephalus No postoperative complications occurred during the study. MRI revealed the progressive dilatation of the cerebral ventricle, gradual narrowing of the subarachnoid space, and consequent thinning of the cerebral cortex consistent with the development of hydrocephalus [Figure - 3]. Evans ratio comparison showed that the difference among the groups was statistically significant (ANOVA, P<0.01). The differences between all paired experimental subgroups was statistically significant (P<0.05) and the differences between experimental subgroups and their corresponding control subgroups was also statistically significant (P<0.05); however, the differences between all of the control subgroups when paired was not statistically significant (P>0.05) [Figure - 2]b. Time-concentration curve The tritiated water of concentration of the chronic subgroup slowly rose to a peak at 16h (only 58% of the chronic control subgroup). The chronic subgroup's tritiated water concentrations were significantly lower than other groups at 1h, 4h, 8h and 16h but higher than other groups at 48h. In other subgroups, tritiated water appeared in the blood at 1h post-injection, then rose to peak at 8h and lowered in magnitude at 48h [Figure - 2]c-e. Statistical analysis of AUC The AUC of the acute or sub-acute test subgroups were slightly higher than their corresponding control subgroups. The chronic test subgroup's AUC was significantly lower than that of its control subgroup. Analysis of variance of the mean AUC showed the differences among the subgroups were scientifically significant (P<0.01).There were no significant differences between all of the paired data sets for the control subgroups, acute subgroup, and sub-acute subgroup (P>0.05), however, the differences between the chronic subgroup and any of the other subgroups was significant (P<0.05) [Figure - 2]f. Discussion Cerebrospinal fluid is absorbed mainly through arachnoid villi [9],[10] and extracranial lymphatic systems. [13],[14],[15],[16],[17] It has been generally accepted by the neuroscience community that CSF absorption through arachnoid villi is the major approach of CSF drainage. CSF may drain through a variety of potential routes including the arachnoid surface [18] and capillary walls. [19] CSF may also be absorbed by the arachnoid membrane adjacent to granulations, but this possibility has not been adequately explored. [18] The elevated CSF pressure accompanying hydrocephalus leads to augmented expression of aquaporin-4 in brain capillaries; [20],[21],[22] however, immediate CSF absorption through brain capillaries has not been observed to date. Animal studies have demonstrated that the lymphatic absorption of CSF accounts for a considerable proportion of clearance. [23],[24] The CSF absorption through the cribriform plate is main point of drainage to the extracranial lymphatic vessels. [25] In our experiment, an external ethmoidectomy was performed to adequately block the lymphatic drainage of CSF through the cribriform plate. The collected data could then indicate the CSF absorptive ability of arachnoid villi clearly. The direct influences of hydrocephalus-inducing agents on the arachnoid villi were avoided in this study. Many previous methods of building hydrocephalus models, including viruses, [26] bacterial inoculations, [27] growth factors such as FGF-1, FGF-2 and TGF-β,[28],[29],[30] neurotoxins, [31] transgenic models of hydrocephalus, [32],[33] and subarachnoid space kaolin injections, [34],[35],[36],[37] affected the function of arachnoid villi in varying degrees. Silicone oil was injected into the fourth ventricle of adult mongrel dogs to induce an obstructive hydrocephalus model in this study. [38],[39] Silicone oil is an "inert" liquid polymer with many properties such as heat-resistance, anti-oxidation, low temperature resistance, radiation resistance, insulation, and hydrophobicity. [40] After the injection of silicone oil into the cerebral ventricle, the silicone oil will gather in the injection position without spreading along the subarachnoid space. It does not lead to inflammation of the ependyma, arachnoid membrane, or cerebral pia mater. [41] Additionally, it does not have any direct effects on the arachnoid villi which are far away from the injection site. The results of MRI and statistical analyses of Evan's ratio confirmed that the obstructive hydrocephalus canine models were successfully induced. The results of this study showed that the tritiated water concentration in the blood of the acute subgroup at 8h was significantly higher than the acute control subgroup. According to the progressive expansion of the ventricular system, we speculated that there must be a significant increase of intracranial pressure (ICP) in the acute phase of obstructive hydrocephalus. [42],[43] Many authors agree that CSF drainage depends on the pressure difference between the subarachnoid space and venous sinus. [44],[45] The significantly higher tritiated water concentration may be related to the abrupt increase of ICP. The tritiated water concentration of the sub-acute subgroup was almost the same to sub-acute control subgroup at 8h. We speculated this was because the CSF absorption of the arachnoid villi was comparatively weaker in the sub-acute phase in contrast to the acute phase of hydrocephalus. Tritiated water concentrations of the chronic test subgroup rose slowly to the peak at 16h (only 58% of the chronic control subgroup). The tritiated water concentrations were significantly lower than other groups at 1h, 4h, 8h and 16h but higher than other groups at 48h. These results indicated that the tritiated water was transported into venous blood via arachnoid villi at a low rate (i.e., CSF absorption of arachnoid villi decreased in chronic hydrocephalus). Other animal experiments have demonstrated subsequent decreases of cerebral blood flow (CBF) and regional cerebral tissue hypoxia when ICP is increased and cerebral perfusion pressure decreased. [46],[47] Our group previously observed the decrease of the plasma membrane vesicles and the increase of Von Willebrand factor levels in arachnoid villi endothelial cells during chronic hydrocephalus. [11] Therefore, we speculated that the decrease of CSF absorption of arachnoid villi in chronic hydrocephalus is related to the damage to arachnoid villi endothelium caused by chronic hypoxic-ischemia. Higher tenascin-C levels and meningeal fibrosis can impair CSF flow through the arachnoid villi in chronic hydrocephalus. [48],[49] Motohashi et al. found sporadic positive cytokeratin staining cells, large numbers of dense extracellular matrices, and leptomeningeal cells deposited in the arachnoid villi in autopsy specimens of patients with chronic hydrocephalus. [50] The core of the arachnoid villi is constructed of dense networks. [51] The protein and cell deposits may obstruct the dense networks of arachnoid villi. Thus, another plausible explanation for the decreased CSF absorption by arachnoid villi in chronic hydrocephalus is the physical obstruction of the dense collagen and elastic fiber networks. The AUC of the acute subgroup and sub-acute subgroup was slightly higher than that of their corresponding control subgroups, but the AUC of chronic subgroup was significantly lower than that of other subgroups. This indicates that the CSF absorption of arachnoid villi has a partial compensatory ability during acute phases and sub-acute phases of hydrocephalus. However, CSF absorption of arachnoid villi was dramatically decreased in chronic phase of hydrocephalus due to possible decompensation. Thus, the disruption of CSF absorption by arachnoid villi is relevant to the duration of hydrocephalus. The longer the state of hydrocephalus, the more severe the damage to the CSF absorptive capacity of arachnoid villi is. Conclusion This study demonstrated CSF absorption by arachnoid villi in canines does not change significantly during acute phases and sub-acute phases of hydrocephalus, but does decrease significantly during the chronic phases. Therefore, the CSF absorption ability of arachnoid villi is greatly compromised in a prolonged state of hydrocephalus. Although the potential mechanism decreases in CSF absorption of arachnoid villi has yet to be elucidated, this study does offer some breakthrough clues into potential therapeutic pathways for obstructive hydrocephalus. This study acts as an advocate for early diagnosis and timely treatment for obstructive hydrocephalus patients. Late shunting operations can relieve physical obstructions of CSF circulation, but the capacity for damaged arachnoid villi to recuperate full functionality may be limited. Acknowledgments We sincerely thank Dr. Adam Paul Allen from the West China School of Medicine, Sichuan University, for his editorial and revision assistance during the preparation of this manuscript. References
Copyright 2010 - Neurology India The following images related to this document are available:Photo images[ni10099t1.jpg] [ni10099e1.jpg] [ni10099f3.jpg] [ni10099f1.jpg] [ni10099f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
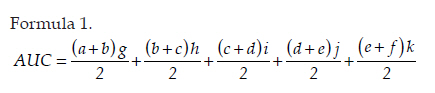{kind=link}
{kind=link}
{kind=link}
{kind=link}