 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
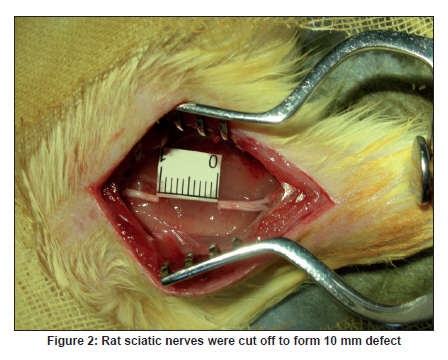
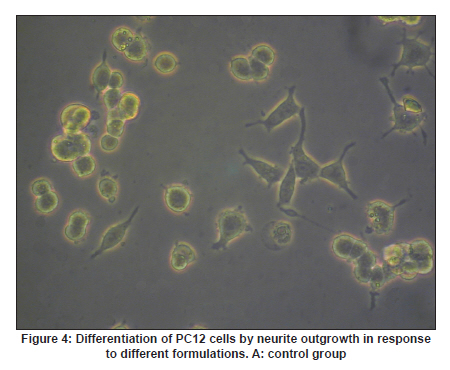
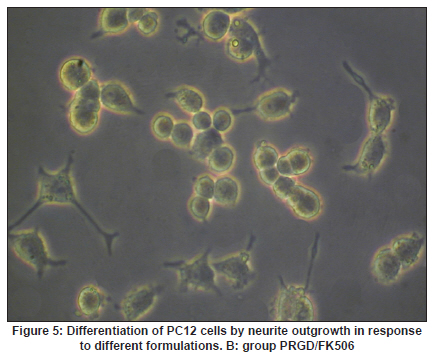
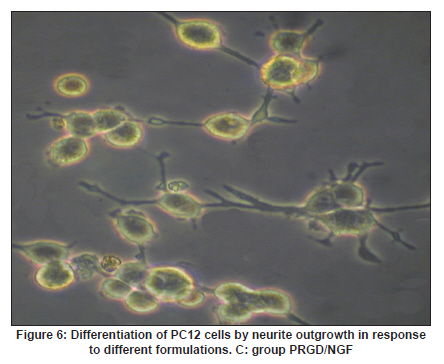
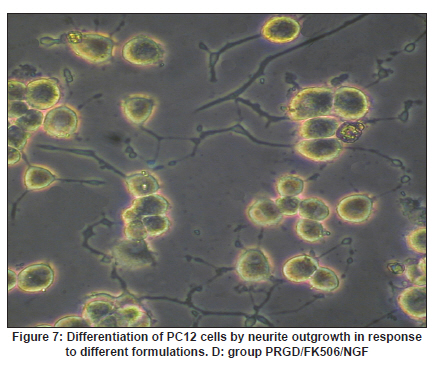
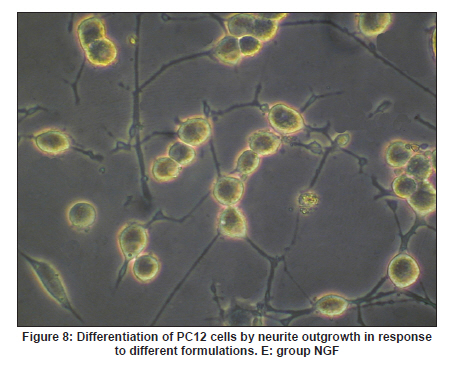
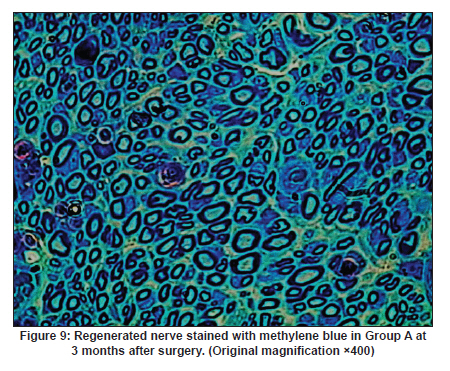
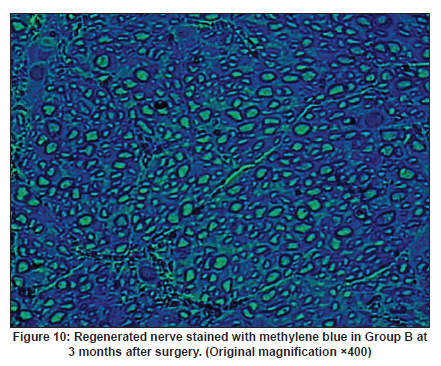
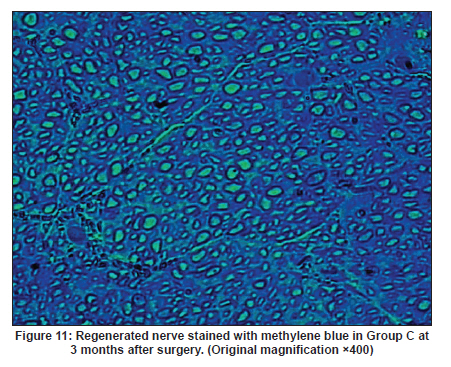
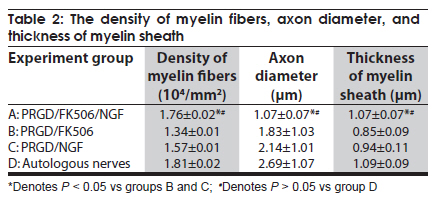
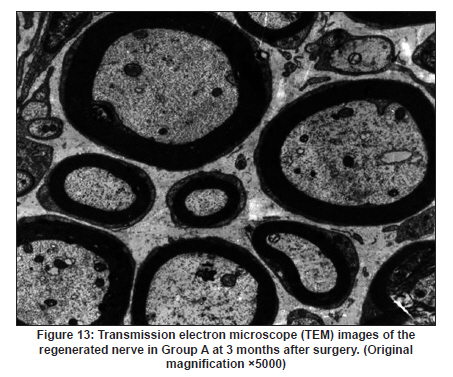
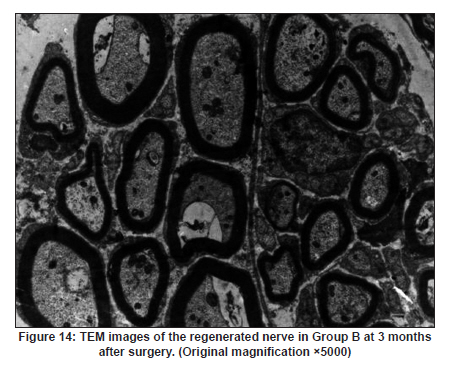
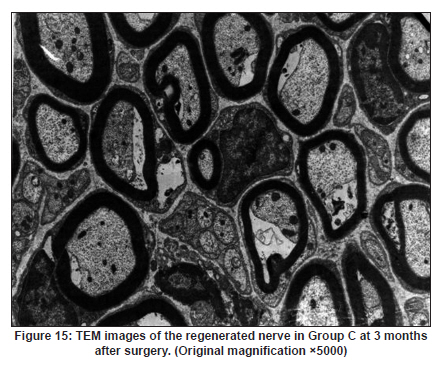
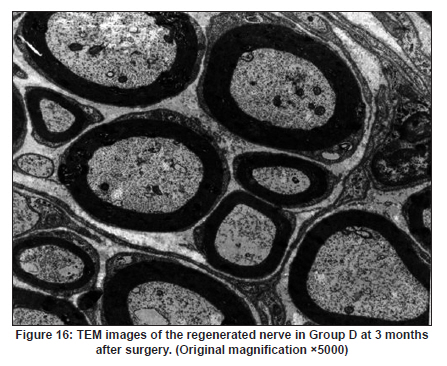
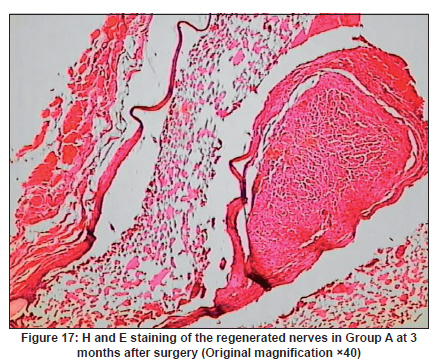
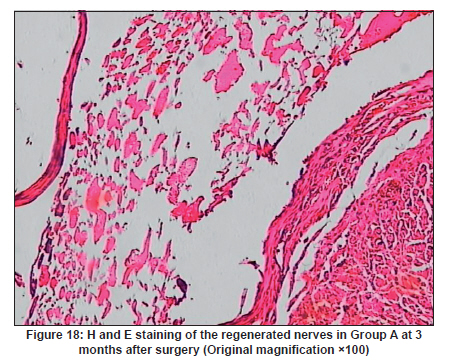
Neurology India, Vol. 58, No. 3, May-June, 2010, pp. 384-391 Original Article Evaluation of PRGD/FK506/NGF conduits for peripheral nerve regeneration in rats Huang Jifeng1, Li Dezhong1, Yan Qiongjiao2, Zheng Huayong1, Li Shipu2 1 Department of Orthopaedics, Wuhan General Hospital of Guangzhou Command, Wuhan, China Date of Acceptance: 11-Jun-2010 Code Number: ni10101 PMID: 20644265 Abstract Context : Both tacrolimus (FK506) and nerve growth factor (NGF) enhance peripheral nerve regeneration, and in vitro experimental results demonstrate that the combination of FK506 and NGF increased neurite outgrowth compared with either treatment alone. Keywords: Nerve conduit, nerve growth factor, peripheral nerve injury, tacrolimus Introduction Peripheral nerve injury is a commonly found clinical phenomenon, but its treatment is far from ideal. [1],[2],[3],[4],[5] Strategies to improve treatment options include treatment with nerve growth factor (NGF) or the use of nerve conduits. [6],[7],[8],[9],[10] Nerve conduits act as a guide for the regenerating axons and as a barrier against the ingrowth of scar-forming tissue. Physical nerve guidance by nerve conduits may not be sufficient to foster optimal recovery. NGF is also required in the treatment of peripheral nerve injury to ensure the survival of the cell bodies and to support the regeneration of the axons toward specific target organs. [11],[12],[13],[14] Although NGF has been shown to improve nerve regeneration under certain conditions, difficulties in the manufacture of these large proteins and delivery to target sites are big barriers to therapeutic application. [15] Tacrolimus (FK506), first used in organ transplantation as an immunosuppressive drug, exhibits neuroprotective and neurotrophic actions in experimental models by increasing neurite elongation and accelerating the rate of nerve regeneration in vitro and in vivo. [16],[17],[18] FK506 is readily synthesized and can cross the blood-brain barrier. In vitro experiments demonstrate that the combination of FK506 and NGF increased neurite outgrowth compared with either treatment alone. [15] To determine if the combination of FK506 and NGF benefits peripheral nerve regeneration compared with either treatment alone in vivo, a novel Gly-Arg-Gly-Asp-Tyr (RGD) peptide modification of poly{(lactic acid)-co-[(glycolic acid)-alt-(l-lysine)]} (PRGD)/FK506/NGF (PRGD/FK506/NGF) sustained-release nerve conduits were fabricated to bridge the 10 mm defect of the sciatic nerve of Wistar rats. Materials and Methods Preparation of the sustained-release nerve conduits PRGD (0.03 g; Biomedical Materials and Engineering Center, Wuhan University of Technology, Wuhan, China) was dissolved in ethyl acetate at a concentration of 10 wt%. FK506 (0.48 mg; Sigma, USA) and NGF (1 μg; Hite Biological Pharmaceutical Co., Ltd., Wuhan, China) were added to ethyl acetate solution and mixed thoroughly. The PRGD sustained-release conduits with FK506 and NGF were prepared using the solvent evaporation method. The same method was applied in both PRGD sustained-release conduits with 0.48 mg FK506 and PRGD sustained-release conduits with 1 μg NGF. The final length of sustained-release conduits was 14 mm, with an inner diameter of 2.0 mm and a tube wall thickness of 0.2 mm [Figure - 1]. The sustained-release nerve conduits were sterilized with ultraviolet light for about 30 min for subsequent experimentation and implantation. Nerve growth factor release In 2 mL of phosphate-buffered saline (PBS) solution (pH 7.4), a PRGD/FK506/NGF conduit was immersed at 37°C. At each time interval, the solution was collected and replaced with fresh PBS. The collected PBS solution was stored at −70°C for later analysis. The NGF in collected PBS solution was quantified using the enzyme-linked immunosorbent assay (ELISA) method, and the bioactivity of NGF in collected PBS solution was examined by culturing PC12 cells. The PRGD/NGF conduits were evaluated using the same method. Cell culture PC12 cells were grown in 25 cm 2 flasks with RMPI 1640 medium (Gibco,USA) supplemented with 10% fetal bovine serum, 5% horse serum, and antibiotics. The cells were cultured at 37°C in a humidified 5% CO 2 atmosphere. Morphologic evaluation and neurite outgrowth assay The experiment was divided into 5 groups (n = 3) (1) PC12 cells cultured in the collected PBS solution with PRGD/FK506/NGF conduits (group PRGD/FK506/NGF); (2) PC12 cells cultured in the collected PBS solution with PRGD/NGF conduits (group PRGD/NGF); (3) PC12 cells cultured in the collected PBS solution with PRGD/FK506 conduits (group PRGD/FK506); (4) PC12 cells cultured in the medium with 50 ng/mL NGF (group NGF); and (5) PC12 cells cultured without NGF supplement as control group. (The collected PBS solution was diluted with culture medium (V/V = 1). PC12 cells were seeded in a 12-well culture plate at a density of 2 Χ 10 4 cells/well, incubated in a humidified incubator at 37°C and 5% CO2. After 7 days, 3-4 random photographs were taken per well, and the neurons bearing processes longer than 5 μm were counted. Both the photographer and the cell counter were blind to the experimental conditions. The neurites were identified and counted from 100 cells per photograph. Animals and surgical procedure Thirty-two Wistar rats weighing 200-250 g each were used and randomized into 4 groups, 8 rats each: Group A: PRGD/FK506/NGF; Group B: PRGD/FK506; Group C: PRGD/NGF; and Group D: autologous nerves. The rats were anesthetized with 40 mg/kg pentobarbital sodium. The right sciatic nerve was exposed after skin incision, and separation of muscles around the nerve tissues was achieved using blunt dissection. Subsequently, the right sciatic nerve was severed into proximal and distal segments in the middle of the right thigh. Defects of 10 mm in the sciatic nerve were created by surgical removal of the nerve tissue. Both the proximal and the distal stumps were secured with 9-0 nylon to a depth of 2 mm into the conduits, leaving a 10 mm gap between the stumps [Figure - 2] and [Figure - 3] in groups A, B, and C. In Group D, the nerve defect was bridged with the resected nerve segment, which was reversed and anastomosed to the proximal and distal nerve stumps. The muscle layer was reapproximated with chromic gut sutures and the skin was closed with silk sutures. All animals were housed in temperature (20°C) and humidity (45%) controlled rooms, and they had access to food and water ad libitum. Electrophysiologic assessment At 3 months after surgery, electrophysiologic tests were performed on all animals. Animals were anesthetized with sodium pentobarbital, the previous surgical site was reopened, and the sciatic nerve was re-exposed. The stimulating electrode was inserted into the proximal neural stem of the regenerated nerve and the recording electrode was inserted into the calf triceps. The peak latency (LAT) and peak amplitude (AMP) of the sciatic nerve action potentials were measured from the chart recordings, as was the conduction velocity through the regenerated nerve. Histologic evaluation The implanting conduits were harvested immediately after the electrophysiologic tests. The regenerated nerves were fixed in 4% glutaraldehyde for 24 h, postfixed in 1% osmium tetroxide for 1 h, and dehydrated in a graded ethanol series. After fixation, some tissues in each group were embedded with olefin and then cut to 4-mm thickness and stained with hematoxylin-eosin (H and E). Other samples were embedded with Epon 812 epoxy resin. The thin-thick slices of 1.0 μm were made and stained with methylene blue. All the nerve sections were observed under a light microscope. HPIAS-1000 high-resolution color graphic report of the pathology analysis system was used to analyze the photographs for the determination of the number of myelinated nerve fibers, myelin thickness, and the diameter of axons of the regenerated nerve. Electron microscopy was performed to evaluate the regenerative nerve ultrastructure. Ultrathin sections of the regenerated nerve tissues were stained with lead citrate and uranylacetate, and then were examined under a transmission electron microscope (Philips CM20, Netherlands). Calf triceps wet weight recovery rate After the implanting conduits were harvested, calf triceps on both the sides were cut out. The blood on the surface of the calf triceps was dried using cotton. Weight of the triceps was immediately measured by electronic weighing scale. Calf triceps wet weight recovery rate (experimental side/normal side) was used to examine the recovery level of calf triceps. Statistical analysis Experimental data were processed using the statistical software SPSS 10.0 (Bizinsight, Beijing China), and expressed as mean standard error of mean. Statistical differences were analyzed using single-factor analysis. The means of the 2 groups were compared using least significant difference law (α = 0.05). Results Nerve growth factor release The amount of NGF released from PRGD/FK506/NGF and PRGD/NGF conduits was analyzed by ELISA and the test was extended for 30 days. At the first day, about 95 ng of NGF was released from PRGD/FK506/NGF and PRGD/NGF conduits. The release rate slowed down thereafter. From day 2 to day 7, on average 19.7 ng/day of NGF was released from the conduits. After day 7, the average minimum daily release of NGF was 4.86 ng. Measurement of neurite outgrowth in PC12 cells The numbers of neurite-bearing cells were calculated with an image processing software. After being cultured for 7 days, about 68.3%, 74.6%, and 10.2% of PC12 cells showed neurite growth in groups PRGD/FK506/NGF, NGF, and PRGD/NGF, respectively. However, less than 5% of PC12 cells showed fiber outgrowth in group PRGD/FK506 and the control group. As shown in [Figure - 4], [Figure - 5], [Figure - 6], [Figure - 7], [Figure - 8], the neurite outgrowth in PC12 cells appeared to show a great difference among the test groups. Electrophysiologic assessment At 3 months after surgery, the compound muscle action potentials (CMAPs) were detected in the hind limbs for each rat in all the groups, and the AMP, LAT, and nerve conduction velocity (NCV) were measured [Table - 1]. The AMP, LAT, and NCV of Group A had a significant difference (P < 0.05) when compared with groups B and C, with no significant difference (P < 0.05) when compared with Group D. The CMAPs of Group A were significantly stronger than those of groups B and C. There was no significant difference between groups A and D. Calf triceps wet weight recovery rate The calf triceps wet weight recovery rates of groups A and D were significantly higher (P < 0.05) than those of groups B and C. The calf triceps wet weight recovery rates were not significantly different between groups A and D (P > 0.05). Histologic assessment The 10 mm gap removed by surgery was found to have been bridged by the regenerated tissue at 3 months after surgery. The regenerative nerve tissues could be clearly observed after opening the conduits. The regenerative nerve samples were removed from the conduits and stained by methylene blue for evaluation in all the groups [Figure - 9], [Figure - 10], [Figure - 11], [Figure - 12]. Numerous bundles of regenerated nerve fibers were clearly found in the sections of the regenerated tissues [Figure - 9], [Figure - 10], [Figure - 11], [Figure - 12]. All the groups were compared for the density of myelin fibers, the axon diameter, and the thickness of the myelin sheath [Table - 2]. The density of myelin fibers, the diameter of the axon, and the thickness of the myelin sheath of groups A and D were significantly greater than those of groups B and C (P < 0.05). The density of myelin fibers, the diameter of the axon, and the thickness of the myelin sheath were not significantly different between groups A and D (P > 0.05). Electron microscopy was performed to evaluate the regenerative nerve ultrastructure [Figure - 13], [Figure - 14], [Figure - 15], [Figure - 16]. Qualitatively, groups A and D appeared to have more organized structure and more uniform fiber shape with larger myelinated fibers than those of groups B and C. There were no significant differences between groups A and D. To evaluate the ingrowth of connective tissues to the wall of conduits, the regenerated nerves with the PRGD/FK506/NGF conduits were stained with HandE. The HandE staining in Group A revealed that no connective tissues were observed to have penetrated into the conduits where nerves should be grown [Figure - 17] and [Figure - 18]. Discussion The curves of NGF released from PRGD/FK506/NGF and PRGD/NGF conduits could be divided into 2 distinct parts each with a different slope, which reflects the 2 different release mechanisms. One was physical adsorption based on electrostatic attraction force NGF and PRGD molecules. In the first part of the curve, the NGF released from PRGD/FK506/NGF and PRGD/NGF conduits showed an initial burst due to the physical adsorption of NGF dissociating from the conduits. This release follows Fick's law. The release behavior will stop within a relatively short time once the concentration gradient disappears. In the second part of the curve the release rate was much slower than the first part. PRGD would be gradually disrupted by hydrolysis and enzymatic degradation. The NGF release behavior follows the degradation-controlled model. Furthermore, the neurite outgrowth assay revealed that there was no obvious neurite outgrowth in PRGD/FK506 group or the control group, a low ratio neurite outgrowth in group PRGD/NGF, and a high ratio neurite outgrowth in groups NGF and PRGD/FK506/NGF. As observed by others, NGF potently stimulated neurite outgrowth with half-maximal stimulation at 1 ng/mL and maximal augmentation at 50-100 ng/mL. FK506 was extremely potent in its neurotrophic effects: with a submaximal concentration of NGF (1 ng/mL), FK506 at 1 nM elicited the same maximal outgrowth observed with NGF at 50 ng/mL. FK506 (100 nM) markedly augmented the effect of NGF by increasing sensitivity to NGF. [15] In the absence of NGF, FK506 failed to elicit neurite outgrowth. The minimum amount of NGF released daily from PRGD/FK506/NGF conduits was about 4.86 ng. Therefore, in the presence of FK506, the daily concentration of NGF released from PRGD/FK506/NGF conduits reached the effective concentration needed to stimulate the maximal neurite outgrowth. Although the peripheral nervous system has a relatively good capacity to regenerate compared with the central nervous system, peripheral nerve repair remains a clinical challenge, as restoration of normal nerve function is highly variable. [19] Nowadays, the clinical technique used for the reconstruction in a peripheral nerve injury is autologous nerve grafts. But this approach has a number of inevitable disadvantages, including extended surgery, loss of donor nerve function, limited supply of donor nerves. [7],[8],[9] To avoid the disadvantages of autografts used in traditional clinical application, many experimental studies have focused on finding alternatives to autologous nerve grafts. As a vital index for the normal function of the peripheral nerve, [20] the CMAPs were used to assess the recovery of the nerve function [Table - 1]. Obvious signals of CMAPs were detected in all the implanted rats, suggesting the quick recovery of nerve conducting function in the implanted rats [Table - 1]. The CMAPs of groups A and D were significantly stronger than those of groups B and C, suggesting a superior conducting function recovery in groups A and D. It is known that the denervation of a target muscle occurs as a consequence of damage to motor nerves. If the muscle is reinnervated, its function is restored and atrophy is stopped. [21] Triceps muscle was taken as a representative of the target muscle of the sciatic nerve in this study. At 3 months after surgery, the calf triceps wet weight recovery rates showed no significant difference between groups A and D, but the calf triceps wet weight recovery rates of groups B and C were significantly lower than those of groups A and D. Histologic assessment showed that the nerve fibers grew better in Group A, and the regenerated nerves had a similar structure compared with Group D. In contrast, groups B and C showed less axon size and myelin thickness. It is well known that axon size and myelin thickness are measures of maturity of the regenerating fibers. Collectively, a better conduction velocity, higher axon counts, and a better recovery of the calf triceps muscle were demonstrated in Group A compared with groups B and C. The results of this study demonstrate that the combination of FK506 and NGF has a positive interaction with respect to peripheral nerve regeneration. Lee et al. [14] reported that the regenerating nerve fibers in the 2 higher NGF dose (20 and 50 ng/mL) groups were not statistically different from the isograft control group. However, the 5 ng/mL NGF delivery system group was statistically different from the isograft control group. Axonal regeneration in the NGF delivery system groups revealed a marked dose-dependent effect in all the assessments. Numerous neurotrophic proteins, such as brain-derived neurotrophic factor and NGF, stimulate nerve regeneration after peripheral nerve injuries. [22],[23],[24] However, difficulties in the manufacture of these large proteins and delivery to target sites are serious barriers to therapeutic application. [15] The objective of this research was to minimize NGF dose and determine if the combination of FK506 and NGF benefits peripheral nerve regeneration compared with either treatment alone in vivo. In previously published reports [15] and in our own observations, the content of FK506-binding protein of PC12 cells can be raised by NGF. In vitro, FK506 has the function to improve the NGF sensitivity of PC12 cells, and to reduce the minimum dosage of NGF by 20-50 times, which has the greatest effect on neurite growth in PC12 cells. However, the biological environment surrounding nerve regeneration in vivo is much more complicated. Therefore, we sought to determine whether the materials we tested in vitro could be successfully applied in vivo. The results of this study demonstrate that the combination of FK506 and NGF benefits peripheral nerve regeneration compared with either treatment alone. Results in all the morphometric measurements demonstrated that a lesser NGF dose, when combined with FK506, was not statistically different from the positive (autograft) control group that provides the clinical "gold standard" for long gap peripheral nerve repair. On H and E staining, no connective tissues were observed to penetrate into the PRGD/FK506/NGF conduits, indicating that the compact structure of PRGD/FK506/NGF conduits well performed its expected functions. The PRGD/FK506/NGF conduits could prevent the connective and scar tissues from growing into the internal wall to disrupt the nerve regeneration. Conclusion The results of this experiment indicate that the combination of FK506 and NGF benefits peripheral nerve regeneration compared with either treatment alone. The PRGD/FK506/NGF nerve conduit promotes axonal regeneration of rat sciatic nerve over a defect of 10 mm, which is close to the success of nerve autograft. These results should encourage further investigation about the potential use of this conduit in clinical study. References
Copyright 2010 - Neurology India The following images related to this document are available:Photo images[ni10101f10.jpg] [ni10101f5.jpg] [ni10101f7.jpg] [ni10101f16.jpg] [ni10101f9.jpg] [ni10101t2.jpg] [ni10101f17.jpg] [ni10101f2.jpg] [ni10101f13.jpg] [ni10101t1.jpg] [ni10101f8.jpg] [ni10101f1.jpg] [ni10101f6.jpg] [ni10101f11.jpg] [ni10101f4.jpg] [ni10101f3.jpg] [ni10101f18.jpg] [ni10101f14.jpg] [ni10101f12.jpg] [ni10101f15.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}