 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
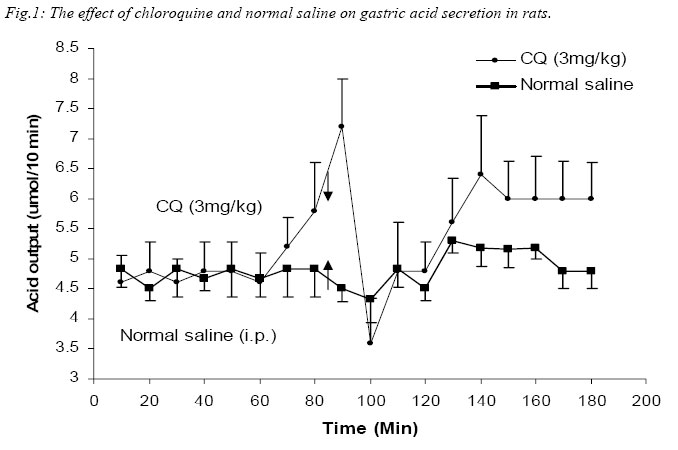
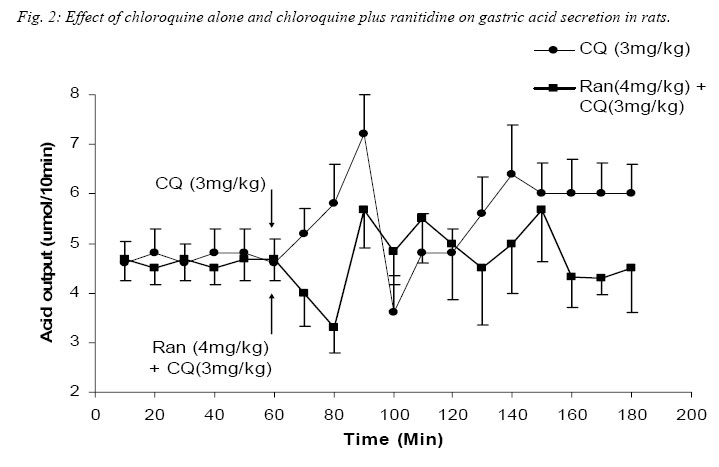
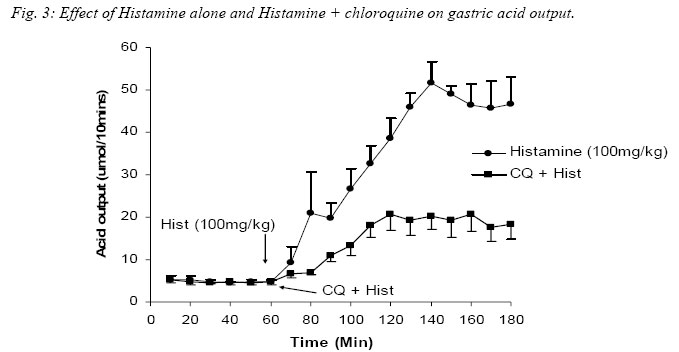
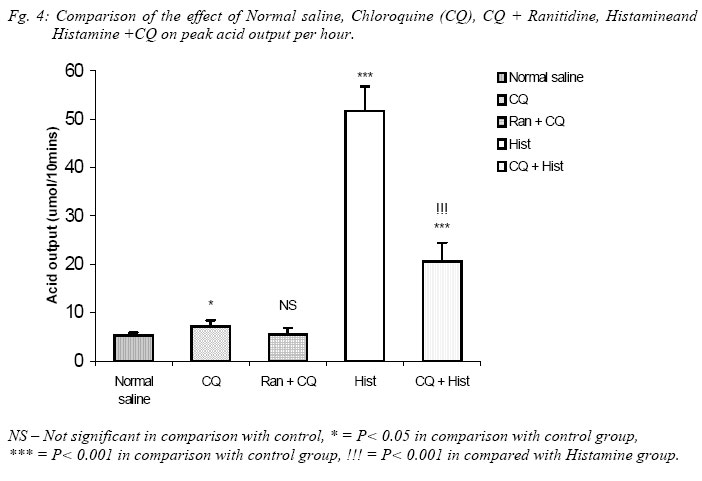
Nigerian Journal of Physiological Sciences, Vol. 20, No. 1-2, 2005, pp. 69-73 PRELIMINARY FINDINGS ON THE EFFECT OF CHLOROQUINE ON GASTRIC ACID SECRETION IN ALBINO RATS. Y. O. ETIMITA, S. A. BISONG, A. B. ANTAI, C. O. NKU, E. E. OSIM* Department of Physiology, University of Calabar, Calabar. Nigeria *Corresponding author Received: 31/10/2005 Code Number: np05013 Summary: It is not certain whether chloroquine-induced pruritus is mainly attributable to the liberation of histamine, a powerful gastric acid secretagogue from mast cells, which may not be beneficial in peptic ulceration. Therefore, the aim of this study was to find out whether chloroquine (CQ) can stimulate gastric acid secretion in the rat. Gastric acid output was measured by the continuous perfusion of rats stomachs under anaesthesia with normal saline at the rate of 1ml per minutes. Thirty albino rats were divided into five groups of six rats each. Three groups had intraperitoneal administration of the following; normal saline (1ml/kg; control), CQ (3mg/kg; test) and Histamine H2 receptor antagonist, Ranitidine (4mg/kg) following CQ administration respectively. The other two groups had subcutaneous administration of histamine (100mg/kg) alone and histamine following CQ administration respectively. The basal acid secretion, (4.71± 0.05μmol/10mins) in a group of rats was not significantly increased in comparison with the peak acid output (p > 0.05) following normal saline administration (1ml/kg; i.p.). Administration of CQ in a second group significantly increased acid secretion to a peak of 7.2 ± 1.7μmol/10mins (p < 0.05). Ranitidine blocked CQ -induced acid secretion in a third group. Histamine significantly increased acid secretion from control level of 4.85 ± 0.14 μmol/10mins to 51.67 ± 5.07μmol/10mins (p < 0.001) in a fourth group, while CQ administered 2mins after histamine administration significantly increased acid level from 4.72 ± 0.12μmol/10mins to peak at 20.63 ± 3.28μmol/10mins (p < 0.001) in a fifth group of rats. The peak acid output in the fifth group was significantly lower than that obtained with histamine alone. In conclusion, chloroquine is a weak stimulant of gastric acid secretion rats. It inhibits histamine-stimulated acid secretion probably by occupying histamine H2-receptors in rats. Key Words: Chloroquine, gastric acid secretion, Histamine Introduction Chloroquine, a 4-aminoquinoline derivative (Clark et al, 1992; Webster, 1992), is commonly used in the chemotherapy of malaria fever, and as an anti-inflammatory disease modifying agent in patients with rheumatoid arthritis or systemic lupus erythematosus (Onigbogi et al, 2000). Chloroquine is highly effective against susceptible strains of plasmodium, producing a radical cure in susceptible faciparum malaria, but suppressive only for Plasmodium vivax and Plasmodium ovale (Clark et al, 1992). The use of anti-histamine following chloroquine administration to suppress chloroquine-induced pruritus experienced by a large proportion of black Africans (Ajayi et al, 1991; Adebayo et al, 1997) suggests that chloroquine stimulates the release of histamine. Evidence to support the release of histamine by chloroquine is in the fact that chloroquine produces venodilation which is mediated by histamine (Abiose et al, 1997). Histamine is a known powerful gastric acid secretagogue and evokes copious secretion of gastric acid from parietal cells by acting on H2 receptors (Garrison, 1992). However, only histamine H1 receptor antagonists are prescribed with chloroquine and no histamine H2 receptor antagonists are given thus suggesting that the histamine released may not affect gastric acid secretion. This suggestion may be unfounded. Therefore, the aim of this study is to find out whether therapeutic dose of chloroquine can stimulate gastric acid secretion in rats and the probable mechanism by which this occurs. Materials and methods Thirty albino rats of Wistar strain weighing between 200 and 300grams were used for the study. The rats were deprived of food 18 hours before the experiment but allowed free access to clean drinking water. The animals were divided into 5 groups, with each group consisting 6 animals. The gastric acid secretion was measured using the continuous perfusion method of Gosh and Schild (1958), modified by Amure and Ginsburg (1964). The animals were anaesthetized with 25% urethane (ethyl carbamate) at a dose 6ml/kg body weight. A tracheal canula was inserted via an incision on the neck to ensure normal breathing through out the course of the experiment. An abdominal incision through the linea alba was made to expose the stomach and a semitransection made at the junction of the pylorus with the duodenum. A pyloric canula was inserted and tied to collect gastric contents. An orogastric canula was inserted for perfusion of pre-warmed (at temperature 370C) 0.9% normal saline (pH 7.00) at a rate of 1ml/minute using a perfusion pump. The animals were kept warm by a 100watts electric lamp to prevent hypothermia. Experimental procedureGastric acid was collect via the pyloric canula at 10 minutes intervals. In order to determine acidity, 10ml of the stomach perfusate was titrated against 0.01M sodium hydroxide (NaOH) solution with phenolphthalein as indicator. Titrable acidity was expressed in micro(µ)mol/10mins. The peak acid output per hour was calculated by multiplying by 6 the rate of acid secretion in the one-sixth hour in which the secretion was the greatest (Davenport, 1997). The test drugs/ substances were used 60 minutes post surgery at what time a steady basal (control) acid secretion has been obtained. Animals in group 1 had normal saline (1ml/kg) injected intraperitoneally (i.p.) after a steady basal was obtained for one hour and the level of gastric acid secretion monitored. The animals in the second group were administered human therapeutic dose of chloroquine (3mg/kg) i.p. (Anigbogu and Badru, 2000) The third group of animals received Ranitidine (4mg/kg i.p.), a histamine H2 receptor blocker, followed by chloroquine. Histamine alone (100mg/kg) was administered subcutaneously (Ganong, 1997) to animals in the fourth group, while animals in the fifth group received histamine followed by chloroquine administration. All animals also served as their own control. Results The effect of chloroquine and normal saline on gastric acid secretion in rats. A steady control (basal) level of acid secretion was obtained for about 60 minutes and afterwards the drugs administered. The mean basal acid output in group 1 (control group administered 1ml/kg normal saline, i.p.) was 4.71 ± 0.05µmol/10mins. On administration of normal saline, a peak acid output of 5.3 ± 0.5µmol/10mins was obtained. These values did not differ significantly. The mean basal acid output in the second group was 4.7 ± 0.47µmol/10mins. Twenty minutes after administration of chloroquine (3mg/kg, i.p.) the acid output rose significantly to a peak of 7.2 ± 1.2µmol/10mins (p<0.05). Even after some fluctuations in acid levels after chloroquine administration, the mean acid output (5.62 ± 0.81µmol/10mins) was still significantly higher than control. Figure 1 shows a comparison between the effect of normal saline (1ml/kg) and chloroquine (3mg/kg) on gastric acid secretion. Effect of chloroquine alone and chloroquine plus ranitidine on gastric acid secretion in rats. In the third group of animals, the administration of CQ (3mg/kg) 2 minutes following ranitidine administration (4mg/kg, i.p.) showed a peak acid output of 5.5 ± 1.33µmol/10mins which was not significantly different from the basal output of 4.61 ± 0.04µmol/10mins. The peak acid output in this group (CQ + Ranitidine), 5.5 ± 1.33µmol/10mins, was also not significantly different from that obtained from the CQ administered group, 7.2 ± 1.2µmol/10mins. Figure 2 compared the effect of CQ alone and CQ and Ranitidine on gastric acid secretion. This did not show any statistically significant difference. Effect of Histamine alone and Histamine + chloroquine on gastric acid output. Histamine(100mg/kg body wt) significantly increased acid output from basal level 4.85 ± 0.14µmol/10mins to a peak of 51.67 ± 5.07µmol/10mins after 80minutes (P<0.001). However, when histamine was administered subcutaneously following intraperitoneal administration of CQ, the peak acid output was 20.67 ± 3.78 µmol/10mins, significantly higher than the steady basal secretion of 4.72 ± 0.12µmol/10mins (P<0.001). The peak acid output was however, lower than (P<0.001) the value obtained with histamine alone. Fig. 3. Comparison of the effect of Normal saline, Chloroquine (CQ), CQ + Ranitidine, Histamine and Histamine + CQ on peak acid output. The peak acid output obtained from CQ (3mg/kg, i.p.) administered group was 7.2 ± 1.2µmol/10mins and it was higher (P< 0.05) when compared to that obtained from the normal saline (1ml/kg, i.p.) administered group, which was 5.3 ± 0.05µmol/10mins (Control). The peak acid output obtained when ranitidine was administered 2minutes before CQ was 5.5 ± 1.33µmol/10mins and it was not significantly different from that obtained for control. When Histamine was administered alone, the peak acid output was 51.67 ± 5.07µmol/10mins, which was significantly higher (P< 0.001) than control and CQ administered group. However, when Histamine was administered with CQ the peak acid output (20.63 ± 3.78µmol/10mins) decreased (P< 0.001) significantly, in comparison with histamine alone although it was still higher (P< 0.001) than control and CQ administered groups (Fig. 4). Discussion Intraperitoneal administration of human therapeutic dose of chloroquine in rats caused significant increase in the gastric acid secretion when compared to the acid output following normal saline administration that was used as control. It would appear that its effect may be due to histamine release since Ezeamuzie et al (1990) and Nosal et al (1991) reported the liberation of low amounts of histamine from isolated rat mast cells by chloroquine. However, another set of our experiments indicated that chloroquine effect on gastric acid secretion may not be due to histamine release. Subcutaneous administration of histamine stimulated a copious amount of acid in the rats which is in consonance with previous reports (Garrison, 1992). However, when histamine was administered subcutaneously following chloroquine administration intraperitoneally in another group of rats, there was also a significant increase in acid output compared with control levels. This increase was however, significantly lower than that obtained when only histamine was given. Chloroquine may therefore not stimulate acid secretion in the rat via histamine release since it did not potentiate histamine-stimulated gastric acid secretion but reduced it. It is likely that chloroquine mechanism on acid secretion may be similar to that of cholecystokinin. Cholecystokinin, a weak gastric acid stimulant, competitively inhibits gastrin-stimulated acid secretion by occupying gastrin receptors and thereby denying them to the more potent hormone (Davenport, 1984). It is likely therefore, that chloroquine weakly stimulates acid secretion but competitively inhibits histamine-stimulated acid secretion by occupying the same histamine H2-receptors. In support of chloroquine occupying Histamine H2-receptors, the administration of ranitidine, histamine H2 – receptor antagonist, two minutes before chloroquine administration resulted in a reduction of acid output comparable to the control level. The reduction in acid output following ranitidine administration suggests that chloroquine weakly stimulated acid secretion in the rats via its action on histamine H2 receptors. There is, however, the possibility of the involvement of other receptors, which were not investigated. In conclusion, chloroquine is a weak stimulant of gastric acid secretion, but it inhibits histamine stimulated acid secretion probably by occupying histamine H2-receptors. If these findings are extrapolated to man, chloroquine may therefore not be beneficial to peptic ulcer-prone individuals. References
© Physiological Society of Nigeria 2005 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}