 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
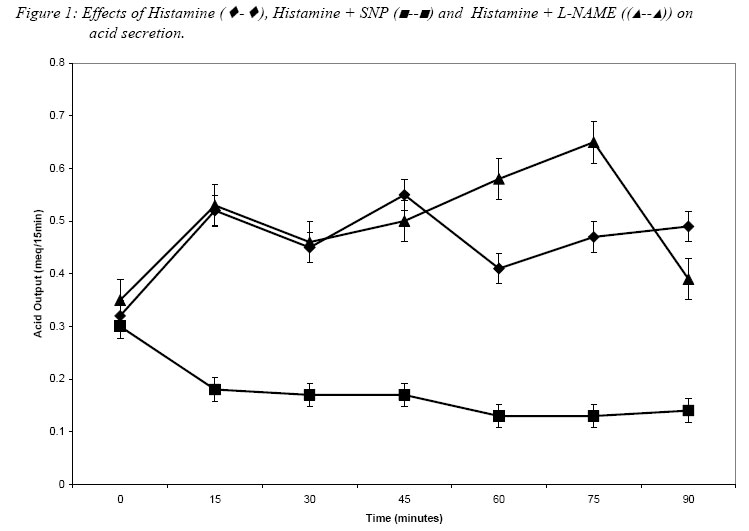
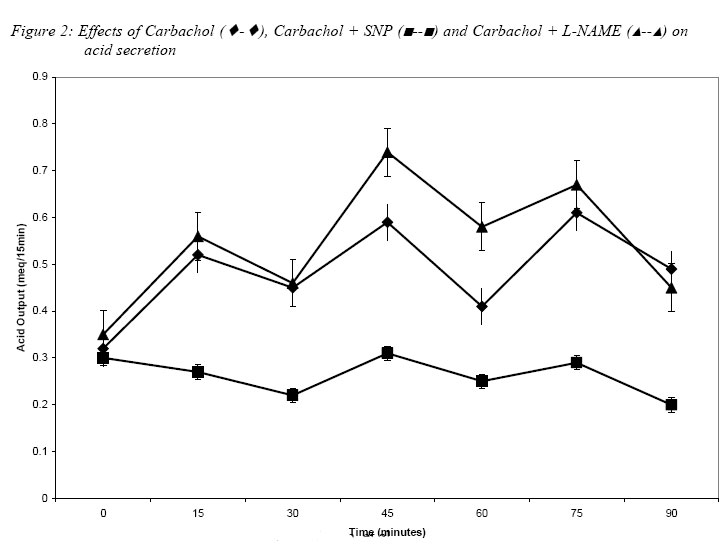
Nigerian Journal of Physiological Sciences, Vol. 20, No. 1-2, 2005, pp. 74-78 INFLUENCE OF NITRIC OXIDE ON HISTAMINE AND CARBACHOL – INDUCED GASTRIC ACID SECRETION IN THE COMMON AFRICAN TOAD – BUFO REGULARIS A. R. A., ALADA, H. M. SALAHDEEN, O.O. AKANDE AND G.O. IDOLOR Department of Physiology, College of Medicine, University of Ibadan, Ibadan – Nigeria Tel./Fax: +234 802 329 1617. E-mail: dralada@yahoo.com Received:24/9/2005 Code Number: np05014 Summary: The study aimed to determine the influence of nitric oxide (NO) on the action of histamine and carbachol on acid secretion in the common African toad – Bufo regularis. Gastric acidity was determined by titration method. The acid secretion was determined when nitric oxide was absent following administration of NO synthase inhibitor; N-nitro-L-arginine methyl ester (L-NAME) and when nitric oxide was in excess by administration of exogenous NO donor, sodium nitroprusside (SNP). Histamine or carbachol increased acid secretion in the toad. Acid output increased from 0.32 ± 0.04 mEq/15min to 0.56 ± 0.08 and 0.61 ± 0.05 mEq/15min for histamine and carbachol respectively (P < 0.05). Pretreatment of the toad with L-NAME produced further increases in histamine (0.62 ± 0.06 mEq/15min) or carbachol (0.74 ± 0.06 mEq/15min) induced acid secretion respectively. SNP however, completely abolished the acid secretion stimulated by either histamine or carbachol. It was therefore concluded that nitric oxide has a negative influence on the histamine or carbachol – stimulated acid secretion in the toad – Bufo regularis. Key Words: Nitric oxide, Histamine, Carbachol, Acid secretion. Introduction In the past one decade, nitric oxide (NO) has been the focus of much interest because of its proposed role in both the central and peripheral nervous system, acting as a diffusible inter- and intra cellular messenger with multifunctional properties (Christopherson and Bredt, 1997; Knowles and Moncada, 1994). In the peripheral nervous system, NO has been reported to regulate gastrointestinal motility, regional blood flow, and neuro-endocrine function. Also, some exocrine and endocrine cells have been shown to possess the NO synthase enzyme which accordingly was assumed to be involved in secretory processes (Ragnar et al. 2002). Gastric acid secretion in mammals and amphibians is regulated by a complex chain of events evoked in response to various nervous and humoral factors. Histamine and carbachol are well-known mammalian acid secretagogues (Oyebola and Alada, 1992). Both drugs have also been shown to stimulate acid secretion albeit in high doses in frogs (Sedar, 1961; Morrisey and Wan, 1970) and toads (Oyebola and Alada, 1992). Histamine stimulates acid secretion through H2 – receptors (Leur et al; 1991), while carbachol acts through the muscarinic cholinergic receptors which are present in the gastric mucosa (Boer et al; 1976). The putative role of NO in the signaling pathway of acid secretion in mammals has been widely disputed and the results obtained are controversial. Pique et al. (1992) showed that NO synthase inhibitor did not affect either the basal or pentagastrin-stimulated acid secretion in rats. Barrachina (1993) reported that NO was a positive modulator of acid secretion in rats. This observation was supported by the report of Sitmewoka (2000) which showed that absence of NO inhibits the secretion of gastric acid. However, Kato et al. (1998) observed that either exogenous or endogenous NO has inhibitory action on gastric acid secretion in mammals. This observation was supported by Takeuchi et al. (1994) who also reported an inhibitory role for NO in the regulation of gastric acid secretion. In contrast, Bilski et al. (1994) reported that NO synthase inhibitor failed to affect basal acid secretion but reduced the acid secretion in response to feeding or pentagastrin in dogs. Much of the controversy however, emanates from the use of different animal models, and application of different qualitative and quantitative techniques in the assessment of acid secretion. Much of the available data on the role of NO in basal and induced acid secretion is on mammals except the work of Molero et al. (1998) who reported that a NO donor L-arginine produced an inhibitory action on basal and carbachol-stimulated acid secretion in toad. Although insensitivity of gastric glands of amphibians to respond to various mammalian secretagogues has been reported (Sedar, 1961; Morrisey and Wan, 1970; Oyebola and Alada, 1992), frogs and toads do secrete gastric acid when the secretagogues are administered in high doses (Oyebola and Alada, 1992). Apart from the study of Molero et al. (1998), there is scarcity of information on the role of NO in gastric acid secretion in amphibians. In view of the scarcity of information and the controversy observed in the results of studies reported for mammals on the role of NO in gastric acid secretion, there is a need to investigate further the influence of NO in gastric acid secretion in amphibians. The present study therefore is designed to investigate the effects of a NO donor- Sodium nitroprusside, SNP and a NO synthase inhibitor- N-nitro-L-arginine methyl ester (L-NAME) on histamine and carbachol induced acid secretion in the Common African toad – Bufo regularis Materials and MethodsExperiments were carried out on seven groups (with ten toads per group) of adult toads weighing 80-120 gm. Toads were collected randomly as described by Oyebola and Elegbe (1975) during the rainy season (May to October) when the toads are commonly found. The toads were fasted overnight and then anaesthetized i.p with sodium pentobarbitone, 3.6 mg/100g. Both the cardiac and the pyloric ends of the toad stomach were cannulated with polythene tubing and the stomach was perfused with 0.7% NaCl at room temperature. Flow rate of the perfusate was adjusted to give an effluent of 1.0±0.1 ml/min. The effluent was collected in aliquots over 15 min per sample. The acid content of each sample was determined by titration with 0.01 N NaOH using phenolphthalein as indicator. In each animal, basal acid secretion was first determined. After the basal collection, the toads in group 1 were injected i.m. with histamine, 3mg/kg and toads in groups 2 and 3 were pretreated with either sodium nitroprusside (SNP), 0.75mg/kg or N-nitro-L-arginine methyl ester (L-NAME – a NOS inhibitor), 10mg/kg i.m. In the pretreatment experiments, about 30 min. was allowed for the drugs to take effect, thereafter, the animals were given an i.m injection of histamine, 3mg/kg.The experiments were repeated on toads in groups 4, 5 and 6 using carbachol, 4.5mg/kg in place of histamine. Toads in group 7 were injected with 0.7% Nacl and served as controls. The effluents from the stomachs were collected at 15 min intervals for 90 min post-injection and the acid content was determined by titration. Acid secretion in all experiments was expressed in mEq/15 min. The mean acid secretion in all the groups studied was calculated and the paired t-test was applied to assess statistical significance of the differences (Bahn, 1972). P values of 0.05 or less were taken as statistically significant. ResultsEffect of Histamine on acid secretion. The effect of histamine on acid secretion in the toad is shown in figure 1. Histamine caused a significant increase in acid secretion with peak acid output of 0.56 ± 0.08 mEq/15 min. This is about 2- fold increase when compared with the basal acid output. Most post- injection values were significantly higher than the basal values. When the animal was pre-treated with L-NAME, the peak acid output increased to 0.62 ± 0.06 mEq/15 min. This effect of L-NAME is most prominent 75 minutes post- injection (figure 1). However, when the toads were pre-treated with a NO donor – SNP, the histamine-induced acid secretion was completely abolished throughout the post-injection observation period. Indeed, the acid secretion fell below the basal level. Effect of Carbachol on acid secretion. The effect of carbachol on acid secretion in the toad is shown in figure 2. Intravenous administration of carbachol also produced significant increase in acid secretion by the toad. The acid output increased from 0.32 ± 0.04 to 0.61 ± 0.05 meq/15min. This increase is about twice that of basal acid secretion. Pretreatment of the animal with NO synthase inhibitor- L-NAME produced further increase in the acid output. The peak acid output increased to 0.74 ± 0.06 mEq/15 min which is an increase of about 60%. Also, when the toad was pre-treated with a NO donor- SNP, there was a complete abolition of the carbachol- stimulated acid secretion. The acid secretion was more or less at the basal level. DiscussionThe significant increase in acid output following administration of histamine or carbachol as observed in the present study is consistent with our earlier report in similar toad experiments (Oyebola and Alada, 1992) and by other workers in other amphibian species (Morrisey and Wan, 1970). Our earlier report (Oyebola and Alada, 1992) had shown that the acid secreting effect of histamine and carbachol is mediated via histamine –H2 receptors. Existence of histamine-H2 receptors in the gastric mucosa of bullfrog has been reported (Watanabe and Goto, 1975). Holstein (1976) suggested that carbachol acts by release of endogenous histamine, which is the final chemo stimulator of acid secretion. Of most interest in this study is that the acid secretory effect of histamine and carbachol were markedly enhanced following inhibition of NO production by L-NAME, the NO synthase inhibitor. The results therefore seem to suggest that the potentiation by L-NAME of histamine and carbachol- stimulated acid secretion most probably results from the removal of inhibitory influence of endogenous NO by inhibiting the production of endogenous NO. This finding is in agreement with the observation of Molero et al (1998) in specie of toad, Bufo bufo, where carbachol and pentagastrin significantly increased NO formation which inhibited acid secretion. The observed inhibition of either histamine or carbachol acid secreting response by a NO donor – SNP in this study is also consistent with the report of Molero et al (1998) where L-arginine (a NO donor) inhibited acid secretion in non-stimulated and carbachol-stimulated gastric mucosa which was reversed by L-NAME, which is a NO synthase inhibitor. Although the literature is sparse on the influence of NO on acid secretion in amphibian species, there are however, several reported studies on the effect of NO on acid secretion in mammals but their results remain controversial (Kitamura et al; 1999). The present results however, clearly showed that endogenous NO exerts a negative influence on acid secretory responses to histamine and carbachol. Since NO synthase inhibitor such as L-NAME is known to cause gastric hypo function (Alumets et al; 1982, Bredt et al; 1990, Esplugues et al; 1996), it is unlikely that the enhanced acid secretion observed under the blockade of NO production is attributable to an increase in the mucosal blood flow. However, Salvemini et al; (1991) reported that exogenous NO inhibited the release of histamine rat mast cell via a guanylate cyclase/ cGMP dependent system. Thus, it was suggested that NO is capable of reducing acid secretion locally by inhibiting the release of histamine from the enterochromaffin-like (ECL) cells (Kitamura et al; 1999). In conclusion, this study has shown that NO has a negative influence on acid secretion induced by histamine or carbachol in common African toad – Bufo regularis. References
© Physiological Society of Nigeria 2005 |

{kind=link}
{kind=link}