 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
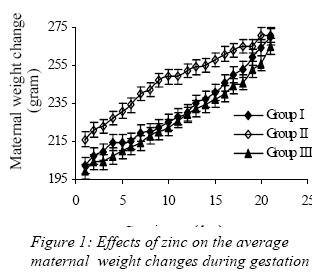
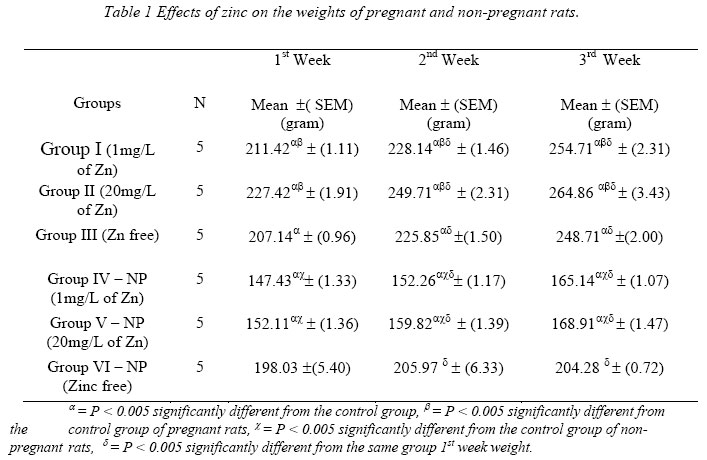
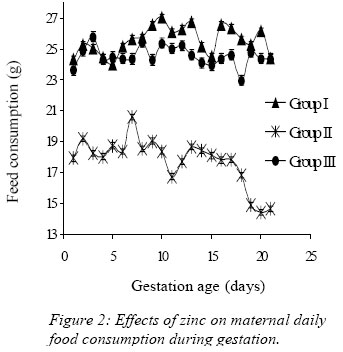
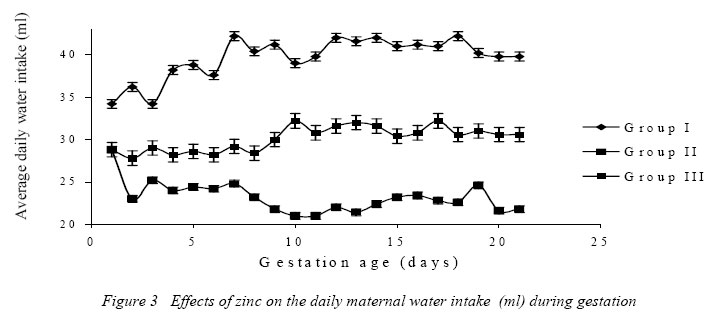
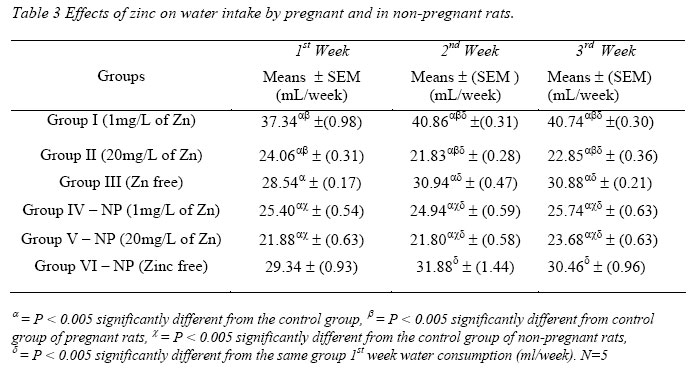
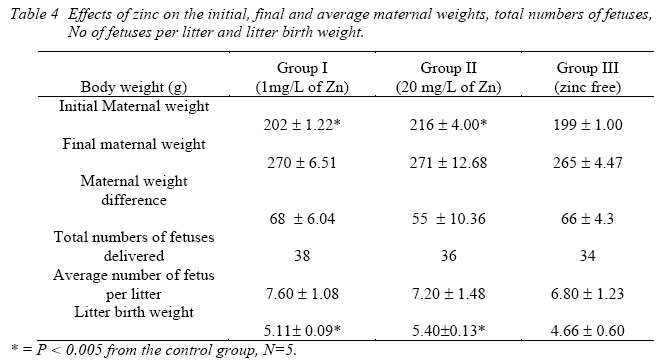
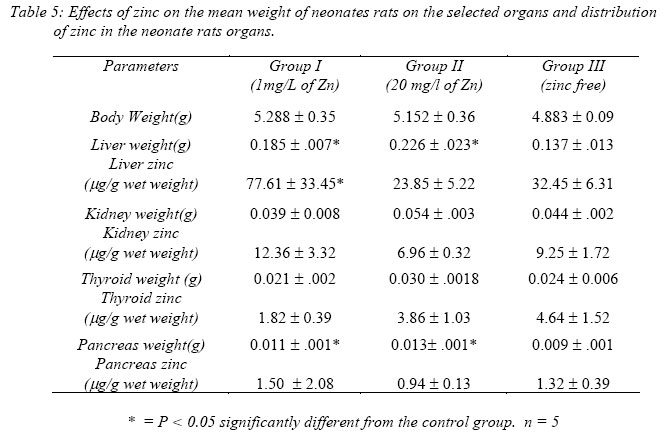
Nigerian Journal of Physiological Sciences, Vol. 21, No. 1-2, 2006, pp. 91-97 SOME EFFECTS OF ZINC ON MATERNAL AND FETAL INTERGRITY IN PREGNANCY. A. A. BADRU, B. I. KUKOYI, and O. E. UKPONMWAN DepartmenCode Number: np06011t of Physiological Sciences, Faculty of Basic Medical Sciences, Obafemi Awolowo University, Ile-Ife Nigeria E-mail: boone@oauife.edu.ng boonekukoyi@yahoo.com Tel: +234 803 344 7197 Received: 15/9/06 Accepted: 12/12/06 Code Number: np06017 Summary: This study was undertaken to determine the effects of zinc sulphate on the weights of pregnant rats, on the food and water consumption during gestation, litters weight at birth, the number of litters at birth and the accumulation of zinc in selected organs of the litters. Zinc sulphate in the dose of 1 mg/liter and 20mg/liter drinking water was administered to both pregnant and non-pregnant for the period of gestation. The results indicated a significant (p<0.005) increase in body weights of pregnant rats when compared with the corresponding controls (non-pregnant and zinc free rats). The values of food consumption in the pregnant rats during 1st, 2nd and 3rd week were significantly (p<0.05) higher than the value obtained for the non-pregnant and the control of non-pregnant rats. Pregnant rats administered with 20 mg/l of zinc drinking water showed a sharp decline in the food consumption from the 3rd week and this was continuous till the end of gestation. The quantity of water consumed by pregnant rats was significantly (p<0.005) higher than the control group (zinc free non-pregnant rats). Non-pregnant rats that received 1 mg/l of zinc drinking water significantly (p<0.005) consumed more water than those that received 20 mg/l zinc drinking water. There was no significant difference between maternal weights of the experimental and control rats at parturition. Rats administered with 1 mg/l zinc delivered the biggest litters-size, which was significantly (p<0.05) different from either those administered with 20mg/l of zinc drinking water or zinc free water respectively. The weights of the liver, thyroid, and pancreas from the litters were significantly (p<0.05) higher than those from the control rats. The liver was found to accumulate significant (p<0.05) higher concentration of zinc when compared with the control group. Key Words: Pregnant rats, litters, and zinc supplement. Introduction Zinc is essential for the growth, development and differentiation of all types of life, including microorganisms, plants and animals (Vallee and Falchuk, 1993; Prasad, 2003). It is essential to the structure and function of myriad proteins, including regulatory, structural and enzymatic (Fredrickson et al, 2003). Zinc is present in all cells and participates in a wide variety of metabolic processes by virtue of its diverse catalytic roles in over 200 enzymes. Zinc enzymes are involved in the synthesis and/or degradation of carbohydrate, lipids, proteins, and nucleic acids (Vallee and Falchuk, 1993). Nutrient needs increase during pregnancy and lactation to support fetal growth and milk synthesis. Therefore, the physiological adjustments that are made to meet those needs must be retained (King, 2000). Demand for trace elements like iron, copper, magnesium, zinc and selenium, are also increased, while their bioavailability to the tissue are altered (King, 2000). Homeorhesis (continual retention of nutrient) occurs when there is retention of the nutrient as observed during growth, reproduction and lactation (Kirchgnesser, 1993). Epidemiological and experimental investigations had revealed that maternal nutrition during pregnancy has a significant effect on fetal development in-utero and later in life during development, and to maturity (Hoet and Hanson, 1999). In rats, maternal under-nutrition in trace elements especially zinc has been attributed to cause delay in growth rate, low birth weight and small sized of most organs (McKenzie et l, 1975). Difficulty in maintenance of pregnancy, stressful parturition, teratogenesis, congenital abnormalities and depressed growth of the offspring are problems that were associated with zinc deficiency during various stage of gestation in rats (Hurley et al, 1971; Warkany and Petering, 1972). The present study was undertaken to determine some effects that are associated with zinc intake in pregnant rats and in the litter response. Materials and MethodsExperimental Animals Thirty adult female Albino rats of Wistar strain, weighing between 150-200 grams were divided into six groups with five rats in each of the group. Three groups served as experimental (pregnant rats) and the remaining three groups (non-pregnant rats) were control. Group I: (n = 5) {Pregnant rats that were given 1mg of Zinc/liter of distilled water orally}. Group II: (n = 5) {Pregnant rats that were given 20mg of Zinc/liter of distilled water orally}. Group III: (n = 5) {Pregnant rats that were given distilled water orally}, Group IV: (n = 5) {Nonpregnant rats that were given 1mg of Zinc/liter of distilled water orally}. Group V: (n = 5) {Non-pregnant rats that were given 20mg of Zinc/liter of distilled water orally}. Group VI: (n = 5) {Non-pregnant rats that were given distilled water orally}. All the animals were housed individually in a metabolic cage and the quantity of food in each are pre-weighed using a sensitive balance. After 24 hours, the food were re-weighed, the difference were recorded as the value of food consumed. 50 ml of oral fluid were given to each rat, and after 24 hours, the difference in volume was recorded as fluid intake. All the animals were fed with feeds containing crude protein: 14.5%; Crude fat: 4.8%; Crude fibre: 7.2%; Crude ash: 8%; Phosphorous: 0.62%. Zinc supplement were administered orally to the entire groups except for the control groups. Zinc sulphate salt dissolve in deionized water served as the source of free zinc. Confirmation of Pregnancy: The mature female Wistar rats were housed together with mature and fertile male Wistar rats fed with the same diet, in a breeding cage over a period of 3-5 days. Vaginal smears were checked daily for confirmation of copulation and were observed under a light microscope for the presence of sperm. The day that the sperm was detected was recorded as day one of gestation for the experimental rats. On 21st day, the nonpregnant rats were sacrificed under chloroform anesthesia. All the animals were weighed and sacrificed. Tissues samples from each individual organ were also weighed. These tissues were digested with 2-ml of concentrated nitric acid and placed in a water bath at 80 oC for 45 minutes to hasten the rate of tissue digestion. Each sample was then diluted with 5-ml zinc free glass distilled water. Zinc was analyzed by a computerized ALPHA-4 Atomic Absorption Spectrophotometer and absorbance was measured at 213.9 nm wavelength. Statistical Analysis Data was statistically analyzed [using both descriptive and inferential] and expressed as mean and standard error of means (SEM). One-way analysis of variance (ANOVA) followed by Student-Newman-Keuls’ test with preset p <0.05 for a significant difference between the mean values when compared with the control (zinc free) rats. Results Figure 1 shows a consistent and progressive linear maternal weight gain in the daily weight of the experimental rats and the control of pregnant rats (group). This graph shows a clear distinction in the weight gain of group II rats, while the group and III later had to get closer toward the end of 3rd week. Group II gained weight in the first 7 days of gestation, while groups I and III start to gain weight from day 10 of gestation. Table 1 shows the comparison in the weekly weights of the pregnant and nonpregnant rats. This table clearly shows a significantly (p < 0.005) increase in body weights of pregnant rats (Group I, II, and III) when compare with the corresponding control groups (non-pregnant and zinc free rats). The average weight (gram/week) in the 3rd week for all of the 6 groups were significantly (p < 0.005) higher than the values observed in the 1st week of the study. Figure 2 shows the effects of different concentration of zinc supplementations on the food consumption in the experimental animals. The pattern of food consumption resembled an irregular cyclical pattern, with food consumption reducing toward the end of each week. Group II rats also show a sharp decline in food consumption from day 15 and this was continuous till the end of gestation. Table 2 shows the comparison in the weekly food consumption in the pregnant and Zinc status in pregnant rats and non-pregnant rats. The values of food consumption for 1st, 2nd and 3rd week for the pregnant rats were significantly higher (p < 0.005) than the values obtained for non- pregnant and control of non-pregnant rats. Within the same treatment group (pregnant in pregnant and non-pregnant rats), comparison between 1st week and other weeks, was found to be significantly (p < 0.005) higher than 2nd or 3rd week except group I and IV. Table 2 further indicated that 3rd week food consumption was significantly (p < 0.005) lower than 2nd week in pregnant rats, but this was not observed in non-pregnant rats except group IV. Figure 3 shows effect of zinc supplementation on water intake during gestation, the figure resembles closely the pattern of a sinusoidal wave, with a tendency of the animals taking much water in alternate days. Although, this was not observed in 3rd week where water intake decrease till parturition. The trend of water consumption in the pregnant rats shows that water intake increased on alternate days and decrease slightly during 1st and 2nd week, but during 3rd week there was an appreciable decrease in water intake. Group II water intake was significantly (p< 0.005) lowers than either group I or group III, this trend was observed from day 1 of gestation till parturition. Table 3 shows the quantity of water intake by pregnant rats (group I, II, & III). The amount of water consumed was significantly (p< 0.005) higher than the control (zinc free non-pregnant rats). Nonpregnant rats that received 1mg/l of zinc supplement, significantly (p<0.005) consumed more water than the 20mg/l of water, but lesser volume when compared with the zinc free non-pregnant rats. Table 4 shows the initial, final, average maternal weight, number of fetus per litter and wet fetal weight. There was no significant (p< 0.05) difference between the maternal weights of experimental and control rats at parturition. Group I (1mg/l of zinc) had a higher value of maternal weight gain but was not significantly different from that of group III or group II. The group II has a low maternal weight gain during gestation. Group I rats delivered the highest number of litters, average number of fetuses for the group I, was not significantly (p<0.05) difference from the other two groups. The zinc-supplemented neonate birth weight was neonatal weight at birth. Group I also had significantly (p < 0.005) higher than the highest number of litters, and higher number control rats. Group II has the highest value of of fetuses per litter. Table 5 shows the effect of maternal zinc supplement during gestation on the weight and the distribution of zinc in the neonate rat organs. The liver, thyroid, and pancreas weight were significantly (p < 0.05) higher than the control rats. The kidney weight however was not significantly (p < 0.05) different from the control rats. Discussion Zinc is an essential trace element and plays important roles as a component of many co-factors and enzymes involved in protein synthesis (Vallee and Goldes, 1984). Clearly, there exists in all cells an optimal zinc requirement for growth and function. There have been numerous and varying reports concerning the involvement of zinc in the growth of cells (Frankers and Telford, 1997). In addition to this fundamental relationship, the effect of zinc on cell growth has been a controversial issue. The present study revealed the effects of exogenous zinc intake on maternal and fetal development during gestation period. It has been reported that zinc deficiency during pregnancy showed a marked effect on fetal growth, and investigations on the mechanism of these effects are complicated by marked changes in appetite and food intake in animals fed with zinc deficient diets (Chester and Quatermain, 1970). Our study did not attempt to produce zinc deficiency in the pregnant rats or in the litters but rather to look into some effects of concentrations of zinc on the pregnant rats with relevance to their weights, the litters at birth and also as it affects the selected organs of neonates. The results obtained indicate that from day one of conception of the pregnant rats to the day 17, group II that was administered with 20 mg/liter of zinc shows a steady increase in body weight when compared with pregnant rats in group I and III. These latter groups subsequently as from day 12 of gestation showed an increase in weight gained, which continued till the end of gestation. This may be due to maternal tissue utilization of zinc, and subsequent accumulation in various soft tissues. Our findings corroborate that of Chester and Will (1973) that the cyclic pattern of feeding as an effective control of food consumption and energy balance in anorexic animals seems as highlighted in Figures 2 and 3. This could not be said to be a classical pattern of food or water intake in anorexic rats but an alternate reduction in food and water intake that met the energy demand of the pregnant rats. There was no weight loss to accompany the reduction in food consumption during the 3rd week of gestation. This observation is in agreement with the proposition of Naismith (1973) that gestation is biphasic in terms of maternal protein or nutrient anabolism and catabolism. The period of catabolism of protein stores and deposition occurred during the 3rd week of gestation causing loss of lean tissue gained in early pregnancy. This is evidently clear from maternal weight pattern of group II in the first 7 days of gestation in rats and reduction in food intake in group II and III in the last 7 days.Further, this study revealed that rats in groups I and IV consumed more water than any other groups, and this could be attributed to either low concentration of zinc in water or the osmotic effects of high concentration of zinc. In group II that received 20 ug/ml of zinc, the concentration of zinc was high and probably affected the taste receptors. Table 4 showed a notably higher value for maternal weight difference in group I and III which can be attributed to the number of fetus delivered at parturition. Whereas, in group II, the significant decrease in the maternal weight could be attributed to reduction in food consumption as observed in their pattern of food consumption. Examination of neonatal weight of the liters indicates a significant effect of zinc supplementation on the birth weights. The group that received the 20 mg/liter of zinc showed the highest weight (Table 4) whilst groups I had a significant (p<0.05) higher birth weights. This study further revealed the relevance of zinc supplementation to the high values of the organs weight recorded for the liver, thyroid and pancreas. However, the weights obtained for the kidney in neonate rats were insignificant. Except for liver of group I neonates (1 mg/liter of zinc), none of the organs accumulate significant amount of zinc in the selected tissues. This confirmed the report of Masuyoshi et al, (1981) that zinc accumulates in liver of animals, and most recently Guan et al, (2003) reported that liver accumulate high levels of zinc which results in altered mitochondrial function. A significantly higher (p<0.05) accumulation of zinc in the liver of neonate rats only suggests that the organ can serve as a storage site for excessive zinc transfer from mother to fetus during gestation. It can be concluded that zinc at low dose for short term may have some beneficial effects on birth weight and also accumulate in the rat liver, the pathological effects of the accumulation in the liver needs to be investigated. References
© Physiological Society of Nigeria 2006 The following images related to this document are available:Photo images[np06017f3.jpg] [np06017f1.jpg] [np06017t4.jpg] [np06017t1.jpg] [np06017t2.jpg] [np06017f2.jpg] [np06017t5.jpg] [np06017t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}