 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 95(2): 151-155, Mar./Apr. 2000 Population Dynamics and Feeding Behavior of Triatoma brasiliensis and Triatoma pseudomaculata, Main Vectors of Chagas Disease in Northeastern Brazil Rodrigo Pedro Pinto Soares/+,
Luciene das Graças Evangelista, Departamento de Parasitologia, Universidade Federal de Minas Gerais, Av.
Antônio Carlos 6627, 31270-901 Belo Horizonte, MG, Brasil *Centro de Pesquisas
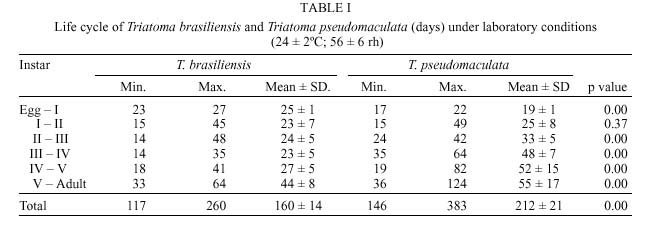
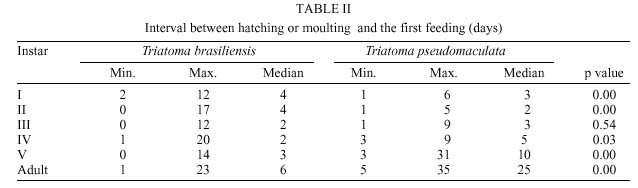
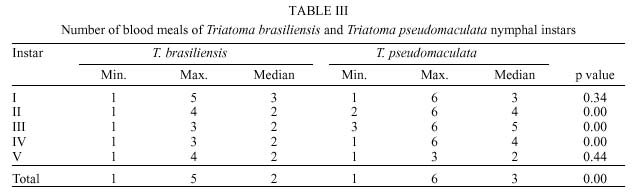
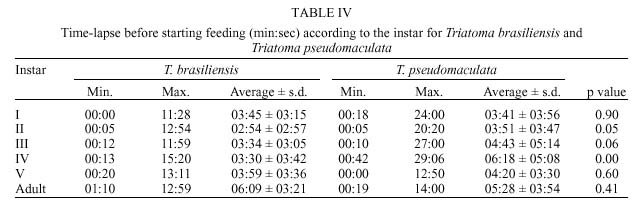
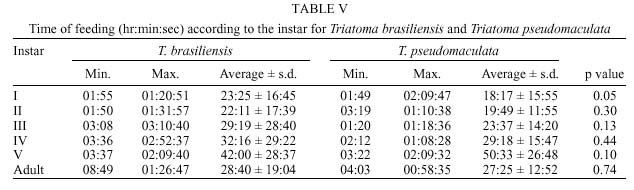
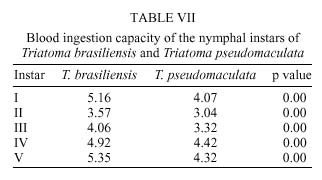
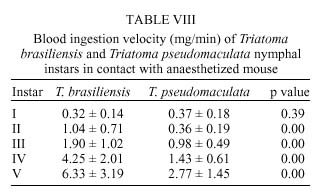
René Rachou-Fiocruz, Av. Augusto de Lima 1715, + Corresponding author. Fax: + 55-31-295.3115. E-mail: rodrigo@cpqrr.fiocruz.brReceived 6 July 1999 Code Number:OC00026 Biological parameters of Triatoma brasiliensis and T. pseudomaculata that could influence the epidemiological importance of these insects as vectors of Trypanosoma cruzi were compared. The parameters studied were incubation period, interval between hatching or moulting and first feeding, number of blood meals, development time, mortality, net reproductive rate, instantaneous daily reproductive rate, time-lapse before starting feeding, duration of feeding, blood ingestion capacity, occurrence of defecation and blood ingestion velocity. Most aspects of feeding were similar for the two species, although T. pseudomaculata had a longer life cycle than T. brasiliensis producing one and two generations per year, respectively. The two species had similar instantaneous daily rates of population growth. Key words: Triatoma brasiliensis - Triatoma pseudomaculata - population dynamics - feeding behavior - Brazil Triatoma brasiliensis Neiva, 1911 and T. pseudomaculata Corrêa & Espínola, 1964 are the most important vectors of Trypanosoma cruzi _ causative agent of Chagas disease _ in the arid caatinga region of Northeastern Brazil. Both species occupy a range of domestic, peridomestic, and sylvatic ecotopes, with T. brasiliensis often associated with small rodents amongst rocky outcrops, and T. pseudomaculata more usually associated with birdnests and hollow trees (Forattini et al. 1981). Application of residual insecticides can eliminate domestic colonies of both species, but houses can then be reinvaded from the silvatic and peridomestic habitats. Our aim in this study, was to compare the two species in terms of various factors that could influence their epidemiological importance as domestic vectors of T. cruzi. MATERIALS AND METHODS Specimens of T. brasiliensis were collected from peridomestic habitats in the municipality of Simplício Mendes, State of Piauí, and specimens of T. pseudomaculata were similarly collected in the municipality of Crateús, State of Ceará, Brazil. Eggs from the collected adults (40 from T. brasiliensis, 33 from T. pseudomaculata) were placed individually in small pots and observed daily until eclosion. The resulting nymphs were offered a daily feed on an anaesthetized mouse. Those that started feeding were allowed to engorge; those that did not start to feed within 30 min were left until the following day. Once bugs had taken a first blood meal, they were then offered a weekly feed. This regime (maintained at 24 ± 2ºC; 56 ± 6% rh) allowed us to establish the incubation period, the interval between hatching or moulting and the first feeding, the number of blood meals required by each instar, the development time, the net reproductive rate (Ro) and the instantaneous daily reproductive rate (r). In the case of 4th and 5th instar nymphs and adults, we also noted the time-lapse before starting feeding, the duration of feeding and timing of defecation. To determine bloodmeal intake, we used 30 bugs of each instar, weighed before and immediately after feeding. The blood meal intake ratios were calculated in relation to the body weight of each insect using the formula (R = (Wf _ Wi)/Wi), where Wf is the weight after feeding (mg) and Wi is the weight before feeding. Blood ingestion velocity was calculated by the formula (V = Wb/T), where Wb is the ingested blood weight (mg) and T is the interval time (minutes) between the beginning and the end of the feeding process. RESULTS Population development - The mean number of eggs laid per female was 264 (range 37-385) for T. pseudomaculata and 142 (range 33-269) for T. brasiliensis. Egg viability was high for both species, with 100% successful eclosion in T. pseudomaculata and 92.5% in T. brasiliensis. Egg development was faster in T. pseudomaculata than in T. brasiliensis, but the duration of instar I to instar II was the similar for both species. Nymphal development during the subsequent instars was longer in T. pseudomaculata than in T. brasiliensis, resulting in an egg-to-adult life cycle of 212 ± 21 days for the former, and 160 ± 14 days for the latter ( Table I). A partial life table analysis, following Southwood (1978), indicates a net reproductive rate (Ro) of 53.2 times per generation for T. brasiliensis and 120 times per generation for T. pseudomaculata under these conditions. The instantaneous daily reproductive rate (r) was 0.010 for T. brasiliensis and 0.009 for T. pseudomaculata ( Table IX). Feeding and defecation - Most aspects of feeding behaviour were similar for the two species ( Table II, Table III, Table IV, Table V, Table VI, Table VII and Table VIII) although nymphal instars of T. brasiliensis tended to be more aggressive than the adults in taking bloodmeals. No differences were apparent in time taken to initiate feeding, nor in the duration of feeding which increased by instar in both species. Some insects fed for more than 2 hr (Table V). T. brasiliensis instars I, III and V had more blood ingestion capacity than T. pseudomaculata. Nevertheless, no difference was observed for instars II and IV (Table VII). All T. brasiliensis instars were faster in obtaining blood meal from the host, except for the instar I (p=0.39) (Table VIII). Defecation during and immediately after feeding occurred in 75%, 86% and 56% of instars IV, V and adults of T. pseudomaculata. For T. brasiliensis the results were 50%, 66% and 50%, respectively (Table VI). Statistical analysis revealed that T. pseudomaculata instars IV and V released feces more frequently than T. brasiliensis instars. No difference was observed for adults. DISCUSSION Epidemiological studies indicate that T. pseudomaculata is less efficient than T. brasiliensis as a vector of T. cruzi. Where both species occur, T. brasiliensis tends to predominate in domestic habitats, with T. pseudomaculata more usually encountered in peridomestic habitats such as chicken coops (Alencar 1987). However, high levels of intradomiciliary colonization by T. pseudomaculata have been observed in the municipality of Sobral, Ceará (Souza et al. 1999), although a survey of 103 houses infested with this species, showed only two people to be seropositive for T. cruzi infection _ indicating a virtual absence of vector-borne transmission (Frota et al. 1999). This contrasts with similar studies of areas infested primarily with T. brasiliensis, for example in the municipality of Independência, Ceará, where studies by our group show a human seroprevalence of 5.7% infection with T. cruzi (Diotaiuti et al. 1998). T. brasiliensis tends to be an aggressive feeder in the field, sometimes attacking people even during the day. Under our experimental conditions however, the aggressiveness of both species was similar. Both species tended to feed for long periods if undisturbed, with some bugs feeding for more than 2 h. However, we noted that T. pseudomaculata was readily disturbed from feeding by even slight movements of the mouse, whereas T. brasiliensis continued to feed even when the mouse made several unconscious movements. Both species defecated frequently while feeding, and T. pseudomaculata instars IV and V defecated more often than the same instars of T. brasiliensis. No such difference was observed for adults. Although the transmission of T. cruzi is crucially dependent on the releasing of feces over the host, the factors involved are complex. Trumper and Gorla (1991) reported that in T. infestans defecation depends on the quantity of blood ingested and on the starvation period. Diotaiuti et al. (1995) showed for T. infestans and T. sordida the occurrence of defecation during and/or immediately after feeding in 90% and 86% of the insects, respectively, compared with our observations of 50% in T. brasiliensis and 75% in T. pseudomaculata. Nevertheless, we suspect that the ready disturbance of T. pseudomaculata could influence its likelihood of completing a feed and defecating on its host. During its development, T. brasiliensis also ingested more blood than T. pseudomaculata, which, in nature, could increase its likelihood of taking an infective blood meal since there seems to be a positive correlation between blood ingestion capacity and the subsequent infection rate by T. cruzi (Minter et al. 1978, Mello & Chiarini 1980). The blood ingestion velocity (mg/min) was the same for instar I of both species, but for the subsequent nymphal instars it was faster in T. brasiliensis, suggesting that T. brasiliensis is more efficient than T. pseudomaculata in obtaining a blood meal. This was reflected in the different development rates of the two species. T. brasiliensis developed faster from egg to adult (160 ± 14 days) than T. pseudomaculata (212 ± 21 days), suggesting that the former would be likely to complete two generations per year, while the latter would be less likely to do so. There were no differences in egg viability, although the incubation period was faster in T. pseudomaculata. For both species the duration from instar I to II was similar, but subsequent development was slower for T. pseudomaculata. Similar results of egg-incubation for T. pseudomaculata were observed by Gonçalves et al. (1997), but for the subsequent instars, our results showed a faster life cycle than that observed by these authors. Several factors may influence these differences, including the length of time adapted to the laboratory, since our observations were made on the offspring of field-collected insects. In addition, our insects were fed on mouse, while those of Gonçalves et al. (1997) were fed on bird (Columba livia). Diotaiuti and Dias (1987) demonstrated that Rhodnius neglectus fed on mouse had a shorter development period compared to those fed on bird. Although T. pseudomaculata seems associated with birds in its natural ecotopes (Forattini et al. 1981), its developmental time seems to be faster when feeding on mouse. The interval between hatching or moulting and the first subsequent feed was shorter for T. brasiliensis, and in general, the number of blood meals before moulting was higher for T. pseudomaculata which sometimes required up to six bloodmeals. For both species, even instar I required more than one feed before moulting, whereas published studies generally show that only a single meal is taken by 1st instars of other species such as R. prolixus (Rabinovich et al. 1979), Panstrongylus megistus, T. infestans and T. sordida (Perlowagora-Szumlewics 1975). There were considerable differences in fecundity between the two species, resulting in marked differences in the net reproductive rates of 53.3 for T. brasiliensis, and 120 for T. pseudomaculata. Because of converse differences in the generation time however, the instantaneous daily rates of population growth were similar, at 0.010 for T. brasiliensis and 0.009 for T. pseudomaculata. These rates are similar to those calculated for T. infestans and R. prolixus (Rabinovich 1972a,b, 1974) under broadly similar laboratory conditions. However the different rates would appear to reflect different environmental adaptations of the two species, in the sense that the greater innate fecundity of T. pseudomaculata may indicate that in nature it may be exposed to higher risk to nymphal survival. This work was supported by CNPq, Centro de Pesquisas René Rachou-Fiocruz and benefited from additional support through the ECLAT network. REFERENCES
Copyright 2000 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc00026f.jpg] [oc00026d.jpg] [oc00026i.jpg] [oc00026h.jpg] [oc00026b.jpg] [oc00026a.jpg] [oc00026c.jpg] [oc00026e.jpg] [oc00026g.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}