 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
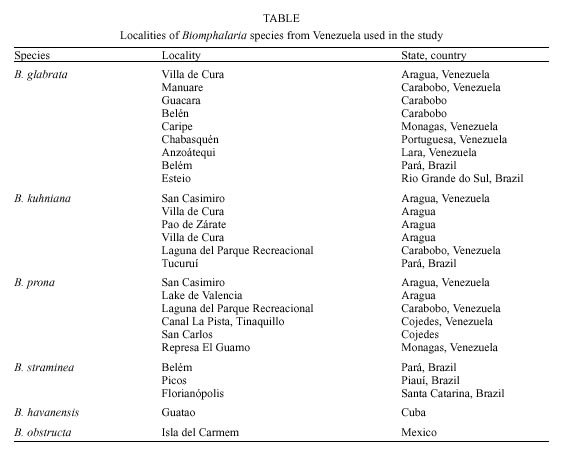
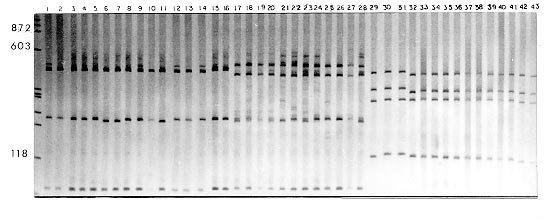
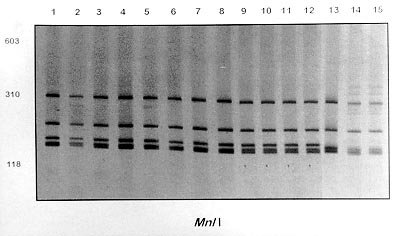
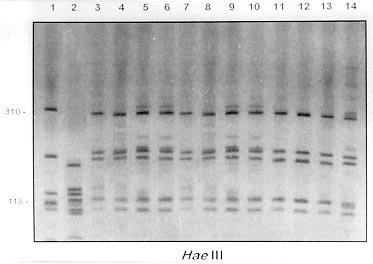
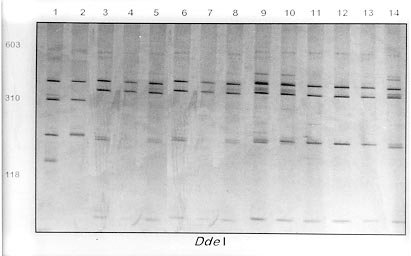
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 95(2): 171-177, Mar./Apr. 2000 Identification of Planorbids from Venezuela by Polymerase Chain Reaction Amplification and Restriction Fragment Length Polymorphism of Internal Transcriber Spacer of the RNA Ribosomal Gene Roberta L Caldeira, Teofânia HDA Vidigal*, Liboria Matinella**, Andrew JG Simpson***, Omar S Carvalho/+ Centro de Pesquisas René Rachou-Fiocruz, Av. Augusto de Lima 1715, 30190-002 Belo Horizonte, MG, Brasil *Instituto de Ciências Biológicas, UFMG, Belo Horizonte, MG, Brasil **Laboratório de Malacologia, Ministerio de Sanidad y Asistencia Social, Aragua, Venezuela ***Laboratório de Genética de Câncer, Instituto Ludwig de Pesquisas sobre o Câncer, São Paulo, SP, Brasil + Corresponding author. Fax: + 55-31-295.3115. E-mail: omar@netra.cpqrr.fiocruz.brReceived 25 August 1999 Code Number:OC00030 Snails of the genus Biomphalaria from Venezuela were subjected to morphological assessment as well as polymerase chain reaction and restriction fragment length polymorphism (PCR-RFLP) analysis. Morphological identification was carried out by comparison of characters of the shell and the male and female reproductive apparatus. The PCR-RFLP involved amplification of the internal spacer region ITS1 and ITS2 of the RNA ribosomal gene and subsequent digestion of this fragment by the restriction enzymes DdeI, MnlI, HaeIII and MspI. The planorbids were compared with snails of the same species and others reported from Venezuela and present in Brazil, Cuba and Mexico. All the enzymes showed a specific profile for each species, that of DdeI being the clearest. The snails were identified as B. glabrata, B. prona and B. kuhniana. Key words: Biomphalaria glabrata - Biomphalaria kuhniana - Biomphalaria prona - snails - internal transcribed spacer - ribosomal DNA The endemic area for schistosomiasis in Venezuela includes the Federal District of Caracas and the states of Aragua, Carabobo, Miranda and Guárico. One third of the country's population lives in this area, where the prevalence of infection with Schistosoma mansoni is 2% (Balzán 1992). To date, seven species of Biomphalaria have been reported in Venezuela: B. glabrata (Martens 1873), B. straminea (Martens 1873), B. prona (Martens 1873), B. kuhniana (Baker 1930), B. havanensis (Hubendick 1961), B. schrammi (Chrosciechowski 1968) and B. obstructa (Chrosciechowski 1988). The first of these species has been shown to be responsible for the transmission of Schistosoma in this country (Balzán 1992). The specific identification of these planorbids is based on comparison of morphological characters of the shell and male and female reproductive organs, as described by Deslandes (1951), Paraense (1975, 1988, 1990), Paraense and Deslandes (1958) and Paraense et al. (1992). Identification is complicated in many cases by morphological variation in these characters, for example when the B. prona population from Lake Valencia (Venezuela), is compared with populations of the same species that occur outside this lake (Paraense et al. 1992). Moreover, identification is also complicated by morphological similarities among some species of snail, for example the B. straminea (Paraense 1988, Caldeira et al. 1998) and B. tenagophila complexes (Spatz et al. 1998, 1999). The polymerase chain reaction amplification and restriction fragment length polymorphism (PCR-RFLP) technique have been used successfully to distinguish several medically important snails of the genera Oncomelania (Hope & McManus 1994), Bulinus (Stothard et al. 1996, 1997) and Biomphalaria (Vidigal et al. 1998a, b, Caldeira et al. 1998, Spatz et al. 1998, 1999). This technique is based on amplification of the internal transcribed spacer region of the rRNA followed by digestion of the resulting fragment with restriction enzymes. This is an easy technique to perform and gives profiles that are simple to analyse. In the present study snails of the genus Biomphalaria obtained from different regions of Venezuela were identified using both morphological and molecular criteria. MATERIALS AND METHODS Snail populations - Snails from different localities of Venezuela were used in the study together with specimens previously identified as B. obstructa from Isla del Carmen, Mexico (type locality); B. havanensis from Havana, Cuba; B. glabrata, B. straminea and B. kuhniana from Brazil. All these species have been reported from Venezuela ( Table). All specimens used were collected in the field, with the exception of B. obstructa from Isla del Carmen which had been maintained in the Departament of Malacology of the Instituto Oswaldo Cruz, Rio de Janeiro and B. havanensis from Guatao, Cuba reared in the Departament of Malacology of the Instituto Pedro Kouri (Havana). All the specimens obtained from the field were examined for cercariae of Schistosoma mansoni and found to be negative. Two individuals from each locality were used for the molecular analysis. Morphological identification of snail populations - Ten specimens of each population were killed, fixed (Deslandes 1951, Paraense 1976) and the feet removed for subsequent DNA extraction. After fixation, each specimen was identified by comparative morphology of the reproductive organs and shell as described by Deslandes (1951), Paraense and Deslandes (1958), Paraense (1975, 1988, 1990) and Paraense et al. (1992). DNA extraction - Total DNA was extracted from the feet of the snails using the Wizard Genomic DNA Purification Kit (Promega) with some modifications. Each foot was mechanically disrupted in 200 µl of nucleic lysis solution and incubated at 37°C for 4 h or overnight with 50 µg/ml of proteinase K, then 80 µl of protein precipitation solution was added to the initial mix. The solution was shaken vigorously and centrifuged at 13,000 rpm for 5 min. The supernatant containing DNA was transferred to a clean tube with 200 µl of isopropanol for DNA precipitation. The mixture was gently homogenized by inversion for 20 min and centrifuged at 13,000 rpm for 5 min. The precipitated DNA was washed with 300 µl of 70% ethanol and centrifuged for 10 min. The pellet treated with 25 µl of DNA rehydration solution for 30 min at 65°C and stored at -20°C. The DNA concentrations were estimated by comparison with known standards on 2% ethidium bromide stained agarose gels. rDNA-ITS amplification - The entire internal transcriber spacer (ITS) (which includes the 5.8S rDNA gene together with the flanking ITS1 and ITS2 spacers) was amplified using the primers ETTS2 (5/-TAACAAGGTTTCCGTAGGTGAA-3/) and ETTS1 (5/-TGCTTAAGTTCAGCGGGT-3/) anchored respectively in the conserved extremities of the 18S and 28S ribosomal genes (Kane & Rollinson 1994). PCR amplification was undertaken in a volume of 10 µl consisting of: 1-10 ng of template DNA, 10 mM Tris-HCl, pH 8.5, 200 µM each dNTP, 1.5 mM MgCl2, 0.8 U of Taq DNA polymerase (Cenbiot, RS, Brazil), 50 mM KCl, together with 1 pmol of each primer. The reactions were covered with a drop of mineral oil and subjected to the following cycle program: initial denaturation step for 5 min at 95°C, and then 32 cycles with: annealing of 54°C for 1 min, extension at 72°C for 2 min, denaturation at 95°C for 45 sec and a final extension step at 72°C for 5 min. A negative control (no template DNA) was included in all experiments. Three microliters of the amplification products were visualized on 0.8% ethidium bromide stained agarose gels to check the quality of amplification. The remaining 7 µl were mixed with 43 µl of water and divided into 10 µl samples for enzyme digestion. RFLP analysis - To evaluate possible enzymes that might yield informative RFLPs of the ITS region, four restriction enzymes (Amersham Life Science) were used: the four base cutters HaeIII and MspI, the five base cutter DdeI and the 11 base cutter MnlI. One microliter (10-12 units) of each enzyme was used for each digestion, together with 1.2 µl of the respective enzyme buffer and 10 µl of the diluted amplification product in a final volume of 12.2 µl. The digestion was performed for 3.5 h at 37°C and products were evaluated on 6% silver stained polyacrylamide gels (Santos et al. 1993, Sanguinetti et al. 1994) after phenol/chloroform extraction to remove protein. The results were recorded with Polaroid film 667 (St Albans, UK). A control for the activity of each enzyme was performed by digesting 150 ng of pUC18 simultaneously with the samples being evaluated. RESULTS Morphological identification of snail populations - The results of morphological identifications of the snails examined are shown in the Table. RFLP analysis - DNA amplification with the primers ETTS1 and ETTS2 generated a fragment of approximately 1.300 bp for all the specimens. Fig. 1 shows the profiles obtained with DdeI enzyme which grouped together: the B. glabrata snails from Brazil and Venezuela (lanes 1 to 16), B. prona populations from the Lake Valencia and from outside this lake (lanes 17 to 28), B. straminea from Brazil (lanes 29 to 31) and B. kuhniana from Venezuela and Brazil (lanes 32 to 43). Figure 2:- silver-stained polycrylamide gels (6%) showing the restriction
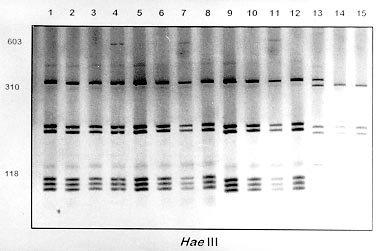
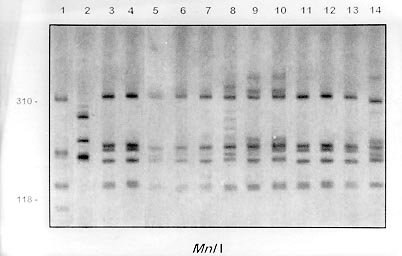
fragment length polymorphism profiles obtained by digest-ing the rDNA internal trancriber
spacer (ITS) with Fig. 2A, B and C shows the profiles obtained with HaeIII, MspI and MnlI respectively. The profiles obtained with theses enzymes grouped together the B. kuhniana from Venezuela and Brazil (lanes 1 to 12) and separated B. straminea from Brazil (lanes 13 to 15). Theses results confirmed the morphological identification and the molecular data showed above with DdeI enzyme. Fig. 3A, B and C shows profiles obtained with HaeIII, DdeI and MnlI, respectively, which demonstrated that theses enzymes permit the separation of B. havanensis (Cuba), B. obstructa (Mexico) of the B. prona populations from Venezuela. Figure 3: silver-stained polycrylamide gels (6%) showing the restriction fragment length polymorphism profiles obtained by digesting of the rDNA internal trancriber spacer (ITS) withHaeIII Fig. 3: (A), DdeI Fig. 3: (B) and MnlI Fig. 3: (C) enzymes. In each gel the snails specimens were: Biomphalaria havanensis: lane 1 Guatao (Cuba). B. obstructa: lane 2 Isla del Carmem (Mexico). B. prona: lanes 3-4 Lake de Valencia, Pan de Azúcar, Club Bahia Paraíso (State of Aragua Venezuela); 5-6 San Casimiro, El Loro Rio (State of Aragua, Venezuela); 7-8 Laguna del Parque Recreacional, Sur de Valencia (State of Carabobo, Venezuela); 9-10 Canal La Pista, Tinaquillo (State of Cojedes, Venezuela); 11-12 Represa El Guamo (State of Monagas, Venezuela); 13-14 San Carlos (State of Cojedes, Venezuela). DISCUSSION The snails B. kuhniana and B. straminea present tenuous morphological differences, making the distinction of these two species the subject of diverse taxonomic problems. Paraense (1988) currently groups them with B. intermedia in the B. straminea complex. Caldeira et al. (1998) were able to separate these two species with the PCR-RFLP technique, with a small genetic distance (0.58) being observed between them in the several populations studied. In the present study, populations from Venezuela previously identified as B. straminea were identified as B. kuhniana based on their morphology and later confirmed by molecular analysis. The data obtained by PCR-RFLP using the enzymes DdeI, HaeIII, MnlI and MspI confirmed the morphological findings. In fact, all the populations exhibited similar profiles to those of B. kuhniana from Tucuruí (Brazil) and were distinct from those of B. straminea from different regions of Brazil. The only reference in the literature to the presence of B. kuhniana in Venezuela was made by Baker in 1930. This record is noteworthy from an epidemiological viewpoint since B. kuhniana is considered to be refractory to S. mansoni (Floch & Fauran 1954) while B. straminea is an intermediate host of this trematode. Paraense and Deslandes (1958), found B. prona (Martens 1873) among material originating from Lake Valencia (Venezuela). Despite the fact that Paraense et al. (1992) later found specimens with morphology and biotypes apparently distinct from the Lake Valencia population in other areas, subsequent morphological comparisons and isoenzyme studies enabled all these snail populations to be identified as B. prona. Although the enzymes used in our PCR-RFLP analysis generated similar profiles for the populations and provide confirmation of this grouping it appears that this species possesses great phenotypic plasticity. The enzymes HaeIII, MspI and MnlI presented complexes profiles, however they were distinct for each species. From the technical point of view PCR-RFLP proved to be effective in the specific identification of these snails, facilitating distinction between Biomphalaria species. ACKNOWLEDGEMENTS To Dr W Lobato Paraense, Departament of Malacology, Instituto Oswaldo Cruz, Rio de Janeiro, for help in the identification of B. kuhniana and for providing B. obstructa. To Dr Gloria Perera Puga, Departament of Malacology of the Pedro Kori Institute, Havana, Cuba, for providing B. havanensis Work partially supported by Fapemig, Pibic/Fiocruz and OPS. REFERENCES
Copyright 2000 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc00030f.jpg] [oc00030b.jpg] [oc00030d.jpg] [oc00030g.jpg] [oc00030e.jpg] [oc00030c.jpg] [oc00030h.jpg] [oc00030a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}