 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
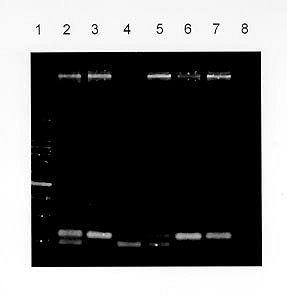
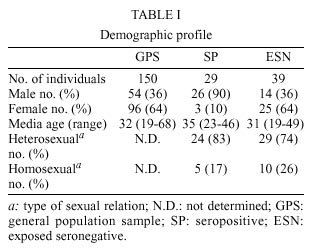
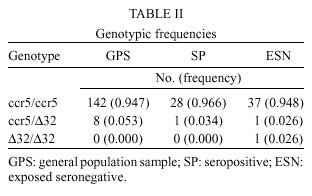
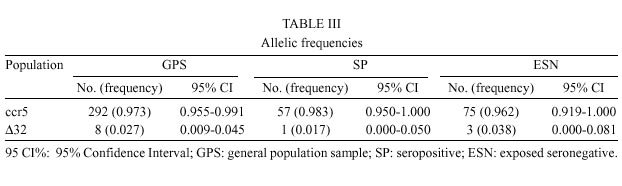
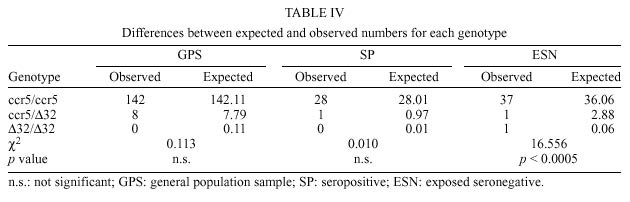
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 95(2): 237-242, Mar./Apr. 2000 Frequency of CCR5 D 32 Mutation in Human Immunodeficiency Virus (HIV)-seropositive and HIV-exposed Seronegative Individuals and in General Population of Medellin, Colombia Francisco J Díaz, Jorge A Vega, Pablo J Patiño*, Gabriel Bedoya, Jorge Nagles**, Cecilia Villegas**, Rodrigo Vesga***, Maria T Rugeles/+ Laboratorio de Virología *Laboratorio de Inmunología, Departamento de Microbiología y Parasitología, Facultad de Medicina, Universidad de Antioquia, AA 1226, Medellin, Colombia **Instituto de los Seguros Sociales, Medellin, Colombia ***Instituto Metropolitano de Salud, Medellin, Colombia + Corresponding author. Fax: + 57-4 510 6062. E-mail: mtrugel@catios.udea.edu.coReceived 26 August 1999 Code Number:OC00041 Repeated exposure to human immunodeficiency virus (HIV) does not always result in seroconversion. Modifications in coreceptors for HIV entrance to target cells are one of the factors that block the infection. We studied the frequency of D 32 mutation in ccr5 gene in Medellin, Colombia. Two hundred and eighteen individuals distributed in three different groups were analyzed for D 32 mutation in ccr5 gene by polymerase chain reaction (PCR): 29 HIV seropositive (SP), 39 exposed seronegative (ESN) and 150 individuals as a general population sample (GPS). The frequency of the D 32 mutant allele was 3.8% for ESN, 2.7% for GPS and 1.7% for SP. Only one homozygous mutant genotype (D 32/D 32) was found among the ESN (2.6%). The heterozygous genotype (ccr5/D 32) was found in eight GPS (5.3%), in one SP (3.4%) and in one ESN (2.6%). The differences in the allelic and genotypic frequencies among the three groups were not statistically significant. A comparison between the expected and the observed genotypic frequencies showed that these frequencies were significantly different for the ESN group, which indirectly suggests a protective effect of the mutant genotype (D 32/D 32). Since this mutant genotype explained the resistance of infection in only one of our ESN persons, different mechanisms of protection must be playing a more important role in this population. Key words: CCR5 - D 32 mutation - human immunodeficiency virus (HIV) infection - exposed seronegative Since the beginning of the acquired immunodeficiency syndrome (Aids) epidemic, it was determined that sexual activity with multiple partners was the main risk factor to acquire human immunodeficiency virus (HIV) infection with the subsequent development of Aids. However, there is strong evidence suggesting a natural resistance to infection in several individuals who have remained uninfected despite the fact that they have had several expositions to HIV, particularly through sexual intercourse (Paxton et al. 1996). The biological basis for this resistance is just starting to be understood. To enter the target cell the HIV requires the presence of the CD4 molecule that acts as receptor and of a second molecule or coreceptor, that in the majority of cases is a chemokine receptor (Deng et al. 1996, Feng et al. 1996). Among the chemokine receptors, CCR5 and CXCR4 are the main coreceptors for HIV entrance. Almost simultaneously with the finding of HIV coreceptors, a mutation in the gene that codifies for the CCR5 molecule was described, which confers a high degree of resistance to HIV infection in vitro and in vivo (Paxton et al. 1996, Liu et al. 1996). This mutation named D 32 consists in a 32 base-pair deletion that encodes for a non-functional protein, and as a result it is not expressed in the cell membrane. Homozygous individuals for this mutation do not have any known immunologic or biologic alteration (Liu et al. 1996). The D 32 allele is present mainly in Caucasian population (Liu et al. 1996, Martinson et al. 1997, Magierowska et al. 1998). In United States the frequency is 8% to 10% in white population but less than 1% in Afro-American individuals. There is also a very low frequency of the mutation among Caucasians in Asia (Pakistan and India) and it has not been reported in China, Japan or pure African population (Martinson et al. 1997). In Latin America the frequencies have been barely studied. It was not detected among 32 individuals from Venezuela nor in Amerindian groups (Liu et al. 1996, Martinson et al. 1997). In Colombia there are no reports addressing this issue. In sexually exposed to HIV but seronegative individuals (ESN), the frequency of D 32/D 32 genotype reaches 2.8% to 3.6%, which corresponds to two to three times the frequency found in Caucasian non-exposed population (Dean et al. 1996, Huang et al. 1996). This frequency can go up to 33% in persons with higher risk of infection (Huang et al. 1996). In contrast, HIV infected individuals exhibit the lower frequency of D 32 allele. It was considered that individuals with homozygous mutant genotype (D 32/D 32) were 100% resistant to HIV infection, at least with M-tropic strains, that uses CCR5 molecule as coreceptor. However, recently there were reports of HIV infection in two individuals who are homozygous for the mutant allele (D 32/D 32) (Biti et al. 1997). These infections could have occurred with T-tropic strains that use the CXCR4 molecule as coreceptor to enter the target cells. It has been difficult to establish if the genotype ccr5/D 32 confers any degree of resistance to infection. The studies on this aspect are contradictory (Samson et al. 1996, Dean et al. 1996, Huang et al. 1996, Hoffman et al. 1997). From the literature one can conclude that D 32/D 32 genotype confers a high degree of protection to HIV infection, but it is not the main factor associated to resistance, since the majority of ESN lack this genetic factor. Different mechanisms such as other coreceptor mutations or immunological factors could account for the lack of infections in this high-risk group. This study was carried out to estimate the frequency of D 32 allele and the genotypes for the ccr5 gene in different subgroups: ESN, seropositive and general population in order to explore the influence of this genetic factor in the behavior of HIV infection in our country. MATERIALS AND METHODS Population -The study was carried out in three groups of individuals. ESN: persons with history of repeated sexual intercourse without protection with HIV infected subjects. The number of subjects in this group is close to the universe of known ESN in our city; SP: all the persons included in this group were sexual partners of ESNs who had antibodies anti-HIV and some of them also presented clinical signs of HIV infection; GPS: this population were adult volunteers, most of them students and workers from our institution or health institutions related to our university. None of them were selected based on risk for HIV infection. The HIV serological status for persons in this group was not determined. Serological status - The serological status for individuals in the ESN group was confirmed by an ELISA test (Enzygnost HIV-1+2 Plus, Behring Diagnostics, Margurg, Germany). Genotype determination - The genotype determination was carried out as previously described (Michael et al. 1997). Briefly, peripheral blood sample was collected with EDTA as anticoagulant. Mononuclear cells were separated by Ficoll-Hypaque gradient. Part of the cells was cryopreserved for future functional studies. The remaining cells were used for DNA extraction using the phenol-chloroform technique. The specific segment of ccr5 gene was amplified by polymerase chain reaction (PCR) using the following primers: CCR5-Delta1 (5'-ACCAGATCTCAAAAAGAAGGT CT-3') and CCR5-D 2 (5'-CATGATGGTGAAG ATAAGCCTCACA-3'). The reaction contained 5 pmol of each primer, 0.3 units of Taq DNA polymerase (Perkin-Elmer), 2 mM of MgCl2 and 200 µM of each dNTP in a final volume of 25 µl. The reaction was subject to 30 cycles of amplification with three temperatures 96°C x 15 sec, 58°C x 60 sec and 72°C x 60 sec. The result of the amplification was electrophoresed in a 2% agarose gel, stained with ethidium bromide and visualized under U.V. transillumination. For wild genotype (ccr5/ccr5) the PCR product was of 225 bp while a product of 193 bp indicated a mutant homozygous (D 32/D 32). Presence of both bands indicated a heterozygous genotype (ccr5/D 32) (Figure). Statistical analysis - The allelic and genotypic frequencies found were compared and the differences evaluated using the chi square test (chi2). The differences between the expected and the observed frequencies of genotypes were evaluated by the same test. RESULTS We studied 218 individuals divided in three groups: ESN, SP and GPS. Their distribution and demographic profile is shown in Table I. All groups had a similar age average. The GPS and ESN groups had a similar distribution of gender but different to SP group in which males were predominant. The majority of couples were heterosexual and few of them were homosexual or individuals non-pared with bisexual preferences. All the seropositive subjects had acquired the infection through sexual intercourse. The genotype frequencies are shown in Table II. The highest frequency was for the wild genotype (ccr5+/ccr5). The heterozygous genotype (ccr5+/D 32) was found in eight GPS, in one SP and in one ESN. Only one homozygous mutant genotype (D 32/D 32) was found among the ESN (Figure). The difference in the frequencies among the three groups studied was not statistically significant. The allelic frequencies for ccr5 and D 32 are shown in Table III. The frequency of the mutant allele D 32 was 3.8% for ESN, 2.7% for GPS and 1.7% for SP. The differences were not statistically significant. Based on the allelic frequencies of each group is possible to predict the genotype frequency considering that they follow Hardy-Weinberg equilibrium. That means that the frequencies have a binomial distribution according to the following equation: p2 + 2pq + q2 = 1, where p and q are the allelic frequencies of ccr5+ and D 32, respectively, and p2, 2pq and q2 are the genotype frequencies of ccr5+/ccr5+, ccr5+/D 32 y D 32/D 32, respectively. A comparison between the expected and the observed genotypic frequencies are shown in Table IV. In SP and GPS the observed frequencies are similar to the ones expected, while in the ESN group the observed frequency differed significantly from the expected frequency getting far away from Hardy-Weinberg equilibrium (p<0.0005). Since the number of individuals in the ESN group and the frequency of D 32 were low, it was not possible to establish a relation between ccr5 genotype and some risk behaviors such as the frequency of the use of condom, type of sexual relation, and time of exposition. The only person with a mutant homozygous genotype was a woman who had a stable sexual relation for three years with a mean of eight non-protected sexual intercourses per month with her infected husband. The subject with heterozygous genotype found in this group corresponds to a promiscuous bisexual man who had frequently anal sexual intercourses during seven months with an infected couple. DISCUSSION The ethnic composition of Colombian population is a product of the mixture of Europeans, Africans and native Indians. Europe contribution is mainly Spanish (Bravo et al. 1996). Two reports of D 32 frequency in Spanish population showed an allele frequency of 8.6% (95%CI 4.9-12.3%) in Basques, 8.2% (95%CI 5.4-10.9%) in Catalans and 5% (95%CI 2.0-8.0%) in individuals from San Sebastian (Martinson et al. 1997, Magierowska et al. 1998). In our study the frequency of D 32 for the GPS group was 2.7% (95%CI 0.90 - 4.5%). Based in this result we could calculate that the Spanish component of the GPS is close or even lower than 50%. This estimation is different from a previous report by Bravo et al. (1996), in which using a panel of different genetic markers they observed a prevalence of the European component over the other races in the population from our province (Antioquia). Since the GPS sample not only included people born in Antioquia but residents of this province, it is possible that a significant number of individuals included in this group were born in different provinces where the influence of the European race is not known. This fact might explain the observed low prevalence of D 32. Although this study found a higher prevalence of D 32 allele and D 32/D 32 genotype among ESN, the differences in the frequencies were not significant (Tables II, III). Probably the reduced sample size did not allow to show a significant association between genotype and infection. In the absence of positive and negative selection or other factors such as frequent recent migration, high mutational rate or high index of endogamy, the distribution of genotypes in each group must be similar to the predicted values following the Hardy-Weinberg distribution. The GPS exhibited a genotypic distribution in equilibrium. Since the circulation of HIV is a selection factor this fact could have two explanations: the frequency of protective genotype D 32/D 32 is too low to be perceptible, or the degree of exposition to HIV is still low in this population. Knowing the frequency of D 32 allele, we calculated the frequency of the D 32/D 32 genotype as 0.00071. This means that only one person among 1406 individuals would carry the mutant homozygous genotype (D 32/D 32). The distribution of genotypes in SP is also according to Hardy-Weinberg equilibrium. The explanation of this fact in this group is more difficult, since all the individuals have been sexually exposed to HIV and this must have positively selected the wild genotype ccr5+/ccr5+. It is possible that the rapid evolution to Aids and the death of individuals carrying wild genotypes over the heterozygous could compensate this effect, but again the low frequency of D 32 allele makes the analysis difficult. The lack of equilibrium in the genotypic frequencies among the ESN was highly significant due to the presence of one individual with the D 32/D 32 genotype. This makes the frequency 17 times superior than expected (Table IV). This finding is hardly explained by one of the following arguments that may also contribute to break Hardy-Weinberg equilibrium in a population: recent European immigration and endogamy, since these facts are not frequently found among our population. The mutational rate for this mutation should not be high either. The possibility that this distribution might have occurred randomly is also very low (p<0.0005). The positive selection of individuals with such genotype D 32/D 32 among ESN is the most plausible explanation for the lack of equilibrium. Therefore, the protective effect of this genotype was shown, although indirectly. In our study the frequency of the D 32/D 32 genotype only explains the resistance to infection in a very low percentage of ESN individuals (2.6%). This is concordant with other studies that have shown similar findings (Bernard et al. 1999). Since many of the individuals in the ESN group have had a high degree of exposition to HIV, there must be other mechanisms responsible for this "natural protection". Recently, it was demonstrated that the HIV could use other chemokine receptors such as CCR2 and CCR3 as correceptors (Berger et al. 1999). Therefore, mutations in these molecules or in the chemokine receptors ligands might explain the resistance to HIV infection of some individuals. However, the mutation CCR2-64I, was not associated with a resistant phenotype but rather with a delayed in disease progression (Smith et al. 1997). Similarly, the polimorphism SDF-1-3'A reported in the alpha-chemokine SDF-1 was associated with the delayed in the onset of Aids related symptoms (Winkler et al. 1998). Other immune system genes such as those of the HLA system appear to also influence the disease progression, although the effects are complex and may depend on interactions with other host genes (McNicholl et al. 1997). Specific cellular immune responses, particularly mediated by cytotoxic T cells or humoral responses at the mucosal level have been demonstrated in exposed seronegative individuals (Mazzoli et al. 1997), indicating that acquired immunity might be the mechanism responsible to control the infection in these individuals. The relative importance of these mechanisms in populations with different modes of exposure or genetic background need to be elucidated This work received financial support from the University of Antioquia and Banco de la Republica. REFERENCES
Copyright 2000 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc00041e.jpg] [oc00041b.jpg] [oc00041d.jpg] [oc00041c.jpg] [oc00041a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}