 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
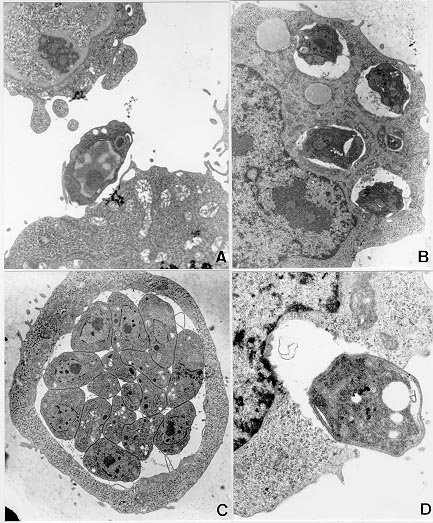
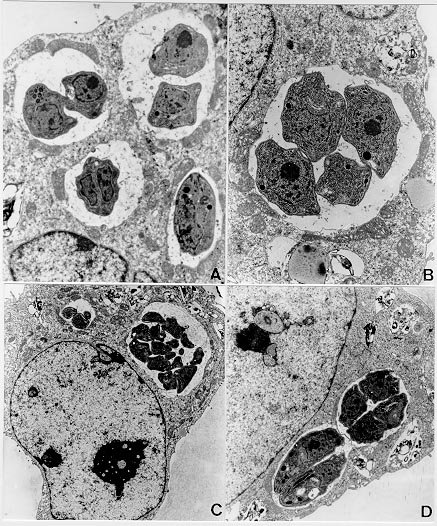
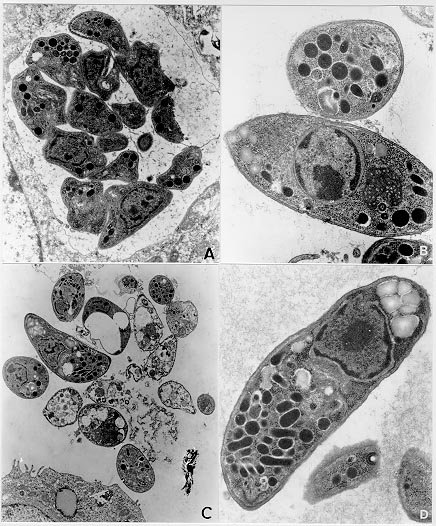
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 95(2): 265-270, Mar./Apr. 2000 Ultrastructural Study of the TG180 Murine Sarcoma Cell Invasion by Toxoplasma gondii: Comparison between in Vivo and in Vitro Cell Cultures Hugo Marcelo Ribeiro Barbosa, Marcos Silva*, Eloisa Amália Vieira Ferro*, José Roberto Mineo/+ Laboratório de Imunologia, Departamento de Patologia *Laboratório de Histologia, Departamento de Morfologia, Universidade Federal de Uberlândia, 38400-902 Uberlândia, MG, Brasil + Corresponding author. Fax: +55-34-218.2333. E-mail: jrmineo@ufu.brReceived 7 June 1999 Code Number:OC00046 Infection of non-adherent TG180 murine sarcoma cells with Toxoplasma gondii was compared, at the ultrastructural level, in both in vivo and in vitro conditions. Suspensions of 3.0 x 106 TG180 cells infected in vitro with 1.0 x 106 parasites of the RH strain were harvested between the first and 6th day post-infection and processed for transmission electron microscopy. In vivo infection was made by intraperitoneal inoculation in mice of 1.0 x 106 TG180 cells, that were co-inoculated with a parasite suspension at the same cell concentration. Cells were harvested 10, 20, 30 min and 24, 48 h post-inoculation and processed for transmission electron microscopy at the same conditions of the in vitro culture. It was observed TG180 murine sarcoma cells with intense and equivalent intracellular parasitism in both conditions. Host cells with parasitophorous vacuoles containing up to 16 parasites, as well as parasites undergoing mitoses or presenting a bradyzoite-like morphology, were frequently seen in both culture methods. Key words: Toxoplasma gondii - TG180 murine sarcoma cells - in vitro culture - transmission electron microscopy Toxoplasma gondii is an intracellular parasite of a wide range of vertebrate hosts, including the human host. The infection caused by this parasite may be asymptomatic in healthy adults, but it can be an important illness to non-immune pregnant women by inducing embryopathy. In immuno-compromised hosts, as in patients suffering of Aids or submitted to transplants, this parasite can originate encephalitis. The most effective treatment currently used is a combination of pyrimethamine and sulphadiaz-ine, which does not eliminate the encysted form of the parasite (Bunetel et al. 1995). Alternatively, it can be used spiramycin, a macrolide antibiotic, which shows no evidence of harmful effects on foetal development or toxicity (Desmonts & Couvreur 1974, Bunetel et al. 1995). There is a low number of toxoplasmicidic drugs on clinical use, but several new drugs have been proposed. New methods are necessary to test these drugs. According to Bunetel et al. (1995), the infection of TG180 murine sarcoma cells in vitro allows an easy, rapid, accurate and economical method to determine the effects of drugs against T. gondii. It has been described in the literature the study of the interaction of T. gondii and host cells by in vivo or in vitro models (Chang et al. 1972, Shepp et al. 1985, Pavesio et al. 1992, Saffer et al. 1992, Speer et al. 1997). The major aim of the present investigation was to study the infection of TG180 cells by T. gondii by comparing cultures submitted to in vivo and in vitro conditions. In addition, it was verified the adequacy of this method, emphasizing the characteristics of the in vitro infection, by using the transmission electron microscopy to evaluate the results. MATERIALS AND METHODS T. gondii - The RH strain used was obtained from animal facilities of our laboratory. The tachyzoites were cultured by intraperitoneal inoculation of female Balb/C mice. The parasites were harvested from mice by rinsing of the peritoneal cavity with 5 ml of Eagle modified minimum essential medium (MEM). TG180 murine sarcoma cell cultures - The TG180 cell line was derived from the ATCC (CCRFS-180 II) sarcoma 180 in 1958 (Sartorelli & Booth 1961). A sub-population of this cell line adapted to in vitro culture (Couatarmanach et al. 1991) was used in our experiments. For the in vitro experiments the cells were grown in MEM supplemented with 10% fetal calf serum, 40 µg/ml gentamycin and 25 µg/ml amphotericin B. The cultures were incubated at 37oC in an atmosphere of 95% air and 5% CO2 in 5 ml-tissue culture flasks. The cells were observed daily by inverted light microscopy, growing in suspension as non-adherent polymorphic cells. Then, 3.0x106 TG180 cells were inoculated into flasks containing 5 ml of MEM culture medium and used after 8-14 passages. Cell countings were made with the use of "Trypan blue". For in vivo experiments, TG180 sarcoma murine cells were grown as ascitic tumors in adult female Balb/C mice in our animal facilities. For the experiments, mice were intraperitoneally inoculated with 1.0x106 TG180 cells. In vitro infection - Six culture flasks were infected with 1.0x106 tachyzoites each. This 3:1 TG180:T. gondii ratio was used as it has been already shown to yield large numbers of intracellular parasites (Couatarmanach et al. 1991). Cell suspensions were collected from the flasks daily from the first to the sixth day post-infection by centrifugation at 1,000g for 10 min. To avoid cell damage due to medium acidification, MEM culture medium was added on the second and third day post-infection. Cell suspensions were fixed overnight with 2.5% glutaraldehyde/2% paraformaldehyde (Karnovsky 1965) diluted in 0.1M phosphate buffer, pH 7.2, washed three times in 5 ml of phosphate buffered saline (PBS) pH 7.4 and processed for transmission electron microscopy as described below. On day 6, culture flasks of uninfected cells were also fixed and used as controls. In vivo infection - Female Balb/C mice were intraperitoneally inoculated with a suspension of 1.0 x 106 TG180 cells. Subsequently, these animals were also inoculated with another cell suspension containing 1.0 x 106 T. gondii by the same route. Mice were sacrificed after 10, 20, 30 min and 24 and 48 h post-infection and their ascitic fluids were collected. This material was fixed with 2.5% glutaraldehyde/2% paraformaldehyde, washed three times in PBS and processed for transmission electron microscopy as described below. Transmission electron microscopy - Briefly, the material was post-fixed for 1 h in 1% OsO4, dehydrated in increasing concentrations of ethanol and embedded in Epon. Ultrathin sections were cut with a diamond knife, stained with uranyl acetate and lead citrate and examined in a Zeiss EM 109 transmission electron microscope. RESULTS Infection of TG180 cells in vivo - The TG180 murine sarcoma cells present a spherical morphology, with finger-like extensions at their periphery after 24 h of infection. In the cytoplasm, it was observed a large amount of free ribosomes and poorly developed rough endoplasmic reticulum. Mitochondria, lipid droplets and multivesicular bodies were also present. The centrally located nucleus is frequently spherical and with chromatin distributed in groups of electron-dense masses. One or two irregularly shaped nucleoli could be observed. After 24 h, T. gondii could be found both in contact with the TG180 cell surfaces and starting the penetration process through invaginations of the host cell plasma membrane (Figs 1a, d). The parasites were also found inside parasitophorous vacuoles after 24 h (Fig. 1b). Vacuoles, free ribosomes, Golgi complex, dense granules, rhoptries and mitochondria of several sizes were present in the intracellular parasites (Fig. 1c). It was also observed TG180 cells with vacuoles containing up to sixteen newly-divided T. gondii in the material collected after 48 h of infection (Fig. 1c). Infection of TG180 cells in vitro - The tumor cells presented similar cellular and subcellular structures as observed in the in vivo condition. We observed the presence of intracellular parasites exhibiting spherical or elongated nuclei with evident nucleoli (Figs 2a, b). Vacuoles, free ribosomes, Golgi complex, dense granules, rhoptries and mitochondria of several sizes were also present (Figs 2b, 2d, 3b, 3c), as observed in in vivo cultured parasites. Parasites undergoing mitotic processes were frequently found in in vitro cultures (Figs 2a, d). There was a low number of intracellular parasites one to two days post-infection and the host cells were seen with single or two parasitophorous vacuoles containing two or four parasites (Figs 2b, d). Furthermore, these vacuoles had their external surface in close relation with mitochondria, which were distributed surrounding them (Fig. 2b). A more accentuated parasitism was seen from three to six days after infection of the tumor cells. After five or six days of infection, it was observed host cells presenting up to eight parasitophorous vacuoles that contained heterogeneous numbers (1 to 16) of parasites (Figs 2c, 3a). These parasitophorous vacuoles had dispersed structures inside them that seemed microtubules (Fig. 3a). We found tachyzoites and cell debris in the extracellular environment coming from the disrupted TG180 cells (Fig. 3c). Parasites morphologically similar to bradyzoites could be also observed (Fig. 3d). DISCUSSION In vitro assays are very important to evaluate the effects of new drugs against intracellular microorganisms, particularly when there are some associated in vivo factors, such as the host immune system, which can interfere in the effects at the intracellular level. T. gondii is an obligatory intracellular parasite that can infect all mammalian cells so far studied. In vitro culture of this parasite can be performed by tissue culture using a broad range of cells (Butenel et al. 1995), including the adherent and non adherent cells. Park et al. (1993) described that under in vitro conditions tumor cells are the most suitable lines for the growth and multiplication of T. gondii. The TG180 murine sarcoma cells are commonly utilized to produce T. gondii by co-inoculation in the peritoneal cavity of mice. Couatarmanach et al. (1991) demonstrated that the culture of TG180 in vitro could produce large amounts of T. gondii, up to 259 parasites/cell. Therefore, this method of culture can be very useful to evaluate the effect of new drugs against T. gondii. We observed by transmission electron microscopy that TG180 murine sarcoma cells are very susceptible to T. gondii infection. Basically, the same parasitism degree was observed in both in vivo and in vitro cultures. We found parasites in the intracellular environments, forming large parasitophorous vacuoles containing up to 16 parasites. Furthermore, extracellular parasites were found close to cellular debris, probably arising from heavily infected cells that released these parasites to the extracellular environment. Ultrastructural analysis of TG180 sarcoma cells showed that these cells, when submitted to in vitro culture, have basically the same cytoarchitectural aspects of the in vivo cultured cells. As a cellular substrate for T. gondii infection, the TG180 can be utilized for diagnostic purpose, or to test new drugs with antitoxoplasmic activity. The TG180 cells can be a good option, as this cell line presents fast growth, is economically viable and is very susceptible to the T. gondii infection. Lindsay et al. (1993) observed in culture of human foreskin fibroblast cells (Hs 68) the presence of host cell mitochondria surrounding the T. gondii tissue cyst wall and suggested that this may contribute directly or indirectly to the formation of the vacuole membrane. Interestingly, we observed frequently the presence of uncountable mitochondria neighboring the parasitophorous vacuoles, some of them completely enclosed by a net of hypertrophied mitochondria, and similar to those described by Lindsay et al. (1993). Another remarkable observation was the presence of parasites in the extracellular environment, some of them harboring an electron-dense substance in their anterior portion that resembled bodies of amilopectine. These parasites presented morphology similar to bradyzoite forms. However, it will be necessary to search for bradyzoite specific antigens in these preparations to confirm these findings. One of the major challenges nowadays is the study of new drugs that can cross the toxoplasmic cyst wall and show toxic effect against parasites located inside these structures. The TG180 murine sarcoma cells may be useful to test new drugs that have their toxoplasmicidic effect just against the bradyzoites. Our results demonstrated that the TG180 murine sarcoma is a suitable cellular substrate to T. gondii culture in both in vivo and in vitro conditions. Thus, this cell type might be used as a potential tool to isolate this parasite with diagnostic aims or as an alternative approach to test new toxoplasmicidic drugs in vitro against both tachyzoites and bradyzoites. ACKNOWLEDGMENTS To Dr Deise Aparecida Oliveira Silva and Dr Maria Aparecida de Souza for their suggestions during the preparation of this manuscript. This work was supported by CNPq, Fapemig and Capes. REFERENCES
Copyright 2000 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc00046a.jpg] [oc00046c.jpg] [oc00046b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}