 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(4) Mar. 2001, pp. 451-458 Simuliid Blackflies (Diptera: Simuliidae) and Ceratopogonid Midges (Diptera: Ceratopogonidae) as Vectors of Mansonella ozzardi (Nematoda: Onchocercidae) in Northern Argentina AJ Shelley/+, S Coscarón* Department of Entomology,
The Natural History Museum, Cromwell Road, London SW7 5BD, UK *Museo La Plata,
La Plata, Argentina Received 23 May 2000.
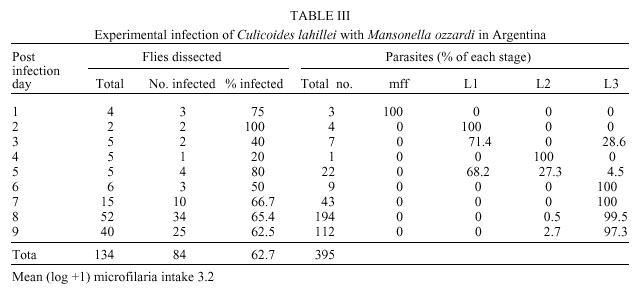
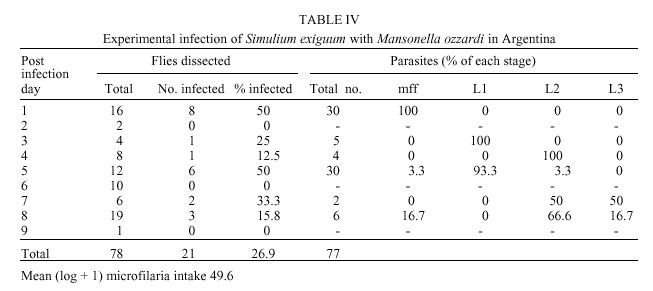
Code Number: oc01069 Mansonella ozzardi, a relatively non pathogenic filarial parasite of man in Latin America, is transmitted by either ceratopogonid midges or simuliid blackflies. In the only known focus of the disease in north-western Argentina the vectors have never been incriminated. This study investigated the potential vectors of M. ozzardi in this area. The only anthropophilic species of these Diptera families biting man at the time of the investigation were Simulium exiguum, S. dinellii, Culicoides lahillei and C. paraensis. Using experimentally infected flies S. exiguum and both species of Culicoides allowed full development of microfilariae to the infective stage, with C. lahillei being a more competent host than S. exiguum. Based on these data, biting rates and natural infectivity rates it is probable that at the begininning of the wet season C. lahillei is the main vector of M. ozzardi and both C. paraensis and S. exiguum secondary vectors. Additionally, it was found that a single dose of ivermectin was ineffectual in eradicating M. ozzardi from infected individuals in this area. Key words: Mansonella ozzardi - Simuliidae - Ceratopogonidae - Simulium exiguum - Simulium dinellii - Culicoides lahillei - Culicoides paraensis - Argentina Mansonella ozzardi is a filarial parasite first described from Guyana (then British Guiana) at the end of the last century and now known to occur in Central America in the Yucatan Peninsula of Mexico through to northern Argentina in South America, as well as on various Caribbean islands. It is relatively non pathogenic, its main importance lying in its morphological similarity to the highly pathogenic Onchocerca volvulus. This is especially relevant in Brazil where the two species are sympatric in parts of the Amazônia onchocerciasis focus. Attempts at controlling mansonelliasis have been few, but recent reports indicate that ivermectin may be effective as a microfilaricide. Various anthropophilic Diptera were examined before it was shown that M. ozzardi is transmitted by Culicoides furens and possibly C. paraensis in the Caribbean Island of St Vincent (reviewed by Buckley 1934, Shelley & Shelley 1976). Later, simuliid species were incriminated as vectors in South America. This gave rise to the hypothesis that two races of M. ozzardi exist: one in the Caribbean islands and Mexico transmitted by ceratopogonid species, and the other transmitted by simuliid species in Panama and South America (Nelson & Pester 1962). Currently, only Culicoides species are known to transmit in the Caribbean islands but both Simulium and Culicoides species are vectors in Central and South America (Table I), but no comprehensive studies have been carried out on the transmission of M. ozzardi and the relative competence of its vectors. Several studies have been made on M. ozzardi in Argentina and these are reviewed in Romaña and Wygodzinsky (1950). Since then several reports have been published on communities in Argentina that are infected with this parasite (Undiano 1971, Jörg 1983, Minoprio et al. 1988, Remon-degui et al. 1988, Taranto & Castelli 1988). These studies indicate that mansonelliasis is confined to the provinces of Tucumán and Jujuy to the north of the country and affects mainly subsistence farming communities, largely formed in the past by the southern dispersal of indians from southern Bolivia. It has been shown that the parasite is relatively non pathogenic, although treatment with ivermectin has been carried out. Attempts at establishing the identity of the vector of M. ozzardi in Argentina have failed. Buckley (1934) and Romaña and Wygodzinsky (1950) cite previous experimental infection studies carried out in Argentina showing that Cimex (Hemiptera, Cimicidae) and Triatoma (Hemiptera, Reduviidae) bugs were not hosts, that microfilariae developed to the L1 stage in Anopheles tarsimaculatus, A. albitarsis and Aedes aegypti but not in A. pseudopunctipennis, A. argyritarsis and C. quinquefasciatus (Diptera, Culicidae). Romaña and Wygodzinsky (1950) showed that no development occurred in simuliids that had fed on M. ozzardi carriers, but that development did occur to the L2 stage in C. paraensis. Development to the L3 stage did not continue because of premature mortality of flies. The objective of the current work was to discover the vector species of M. ozzardi in the mansonelliasis focus in northern Argentina. MATERIALS AND METHODS Study area - The work was carried out in January 1998 at the beginning of the wet season in subtropical cloud forest (Yungas) in the mountain range Quebrada de Humahuaca, to the east of the Andean foothills and north of San Salvador de Jujuy in Jujuy Province in north-west Argentina. The sparse population (300) in this area, drained by the River Candelaria and its tributaries, is involved in subsistence agriculture and timber extraction; in the lower lying areas nearer the town of Fraile Pintado orange groves and sugar plantations provide the main agricultural activities. Laboratory facilities were used in the hospital in the town of Fraile Pintado and the school at Aforo (previously Arrayanal, 23°54'S65°02'W, altitude 900 m), some 50 km to the west of this town on the confluence of the River Candelaria and Arroyo Normenta, was used as a base for field work. This was centred on riverine communities (called Finca Normenta) of mainly local subsistence farmers living farther up the Arroyo Normenta. Selection of volunteers - Remondegui et al. (1988) carried out a survey of mansonelliasis and its potential vectors in the Finca Normenta area ten years ago and found prevalence rates in villages varying between 25-75%. In November 1995 a survey was made in the same area by staff from the hospital at Fraile Pintado to register the names of individuals infected with M. ozzardi, followed in December 1995 by a single treatment of infected individuals using ivermectin (pers. comm. Dr G Safarov). This register of individuals infected with mansonelliasis with information on their treatment was used as a basis for selecting people for the present study. A lancet was used to puncture the fourth finger of each individual and a drop of blood used to make a thick film on a microscope slide. The film was de-haemaglobinized, fixed in 80% methanol, stained with Giemsa and then examined for microfilariae. Prospection for simuliid and ceratopogonid species - Fifty years ago Romaña and Wygodzinsky (1950) surveyed an area in Tucumán Province to the north of our study area and considered the following anthropophilic species to be potential vectors of M. ozzardi: S. dinellii, S. jujuyense, C. paraensis and C. lahillei (as C. debilipalpis - now considered a junior synonym of C. lahillei). Previous prospection of the Finca Normenta area by Remondegui et al. (1988) in the dry season (pers. comm. Dr G Safarov) revealed the presence of three simuliid species (two unnamed) of which S. dinellii was predominant and 16 species of ceratopogonids (all unnamed). In the present survey immature stages of Simuliidae were collected from the rivers Candelaria and Normenta for several hours on several days at the beginning of the study. Larvae and immature pupae were preserved in 80% ethanol, as were adults that emerged from mature pupae kept individually in tubes containing moistened paper. Linked reared adults were also micropinned and, together with their pupal exuviae, preserved in a small phial of glycerine attached to staging pins. Adult Simuliidae and Ceratopogonidae were also collected for short periods over several days biting ourselves, horses and cattle in order to establish which were the anthropophilic species in the area. Specimens of C. lahillei were later dissected for filariae. Identification of simuliid species was carried out by Prof. S Coscarón and the ceratopogonid species by Dr G Spinelli of the La Plata Museum. Voucher specimens have been deposited in the La Plata Museum and the Natural History Museum, London. Infection of flies - In order to establish the hosts of M. ozzardi samples of local biting flies were experimentally fed with the parasite to observe any subsequent development and other flies were dissected for natural infections with filariae. The volunteer used for experimental infection studies was a highly infected 74 years old male, who has lived in the area all his life. He was positive in the 1995 survey, was treated with ivermectin then, but was still highly positive in the present survey. Simuliids and ceratopogonids were allowed to feed to repletion on the volunteer's legs and a small proportion of these flies had their abdomens removed and the blood in their stomachs smeared on a microscope slide for counting and identification of microfilariae using the technique given above for thick films. The rest were maintained alive for up to nine days in individual tubes containing filter paper and a cotton wool wick soaked in 20% boiled sucrose solution. Tubes were kept in the dark in an insulated container in which the temperature varied from 21-30°C. The tubes were examined daily for dead flies, which were stored in 80% ethanol and all living flies nine days after experimental infection were sacrificed and similarly stored. The cotton wool wicks were changed every two days. Preserved flies were later examined in the laboratory for filariae using the following method. Flies were removed from ethanol and placed in tap water over night for re-hydration. Each fly was then placed in a drop of tap water and divided into head, thorax and abdomen. These parts were then added to a drop of Mayer's haemalum for 30 min, transferred to a drop of tap water to remove excess stain and then placed separately in three drops of glycerine on a microscope slide. The head, thorax and abdomen were then dissected and parasites removed from each site were mounted in Berlese mountant for identification using morphological characters described in Shelley et al. (1980) and Tidwell et al. (1980). Collections of simuliids and ceratopogonids were also made from human volunteers to establish whether any natural filarial infection was present. These flies were immediately preserved in 80% ethanol following capture, and were later processed and examined in the laboratory using the method detailed under experimental infection. RESULTS Prevalence of mansonelliasis - In order to select a volunteer with a high microfilaraemia for the experimental infection work, individuals, who were known to be positive in the 1995 survey, were re-examined, even if they had been treated with ivermectin. Table II shows the results of this survey. Although our blood sampling was not done quantitatively the results clearly show that despite ivermectin treatment four individuals out of the 11 treated were infected with M. ozzardi. Simuliid and ceratopogonid species - The following species of Simulium were recorded in the area: S. exiguum Roubaud, S. dinellii (Joan), S. romanai Wygodzinsky and S. wolffhuegeli (Enderlein). Small numbers of S. exiguum and S. woffhuegeli were collected biting the ears and belly of horses, but only the occasional S. dinellii and moderate numbers of S.exiguum bit man. The Ceratopogonidae recorded from the area were: C. paraensis (Goeldi) and Forcipomyia stylifera Lutz biting horses in low numbers, C. lahillei (Iches) biting cattle in low numbers and C. lahillei (263) and C. paraensis (46) biting man. Infection of flies - Only C. lahillei and S. exiguum were collected in large enough numbers to analyse the development of M. ozzardi. There was a considerable difference in mean ingestion rates of the two species that could not be explained by the small size of the former species relative to S. exiguum. C. lahillei only ingested 3.2 microfilariae [mean (log + 1)] per fly (range 0-17, n=14) compared to 49.6 [mean (log+1)] (range 15-186, n= 12) in S. exiguum. A single S. dinellii collected replete from the bait contained 42 microfilariae. Table III gives details of the experimental infection of C. lahillei with M. ozzardi. Of 134 flies experimentally infected 62.7% (84) later showed 395 developing filariae. L3 larvae were first found on post infection day (PID) 3, probably resulting from a natural infection in these wild caught flies as is possibly the case on PID 5. The largest numbers of L3 larvae found were 14 in the thorax and eight in the head. The majority of parasites had reached the final stage in samples from PID 6-9 showing that it has a synchronous cycle.The mean (log+1) production of L3 larvae during PIDs 6-9 was 1.7, signifying that 53.1% of the 3.2 microfilariae ingested per fly developed to the infective stage. Fewer microfilariae ingested by S. exiguum developed in the fly (Table IV) despite a 15 times higher intake of parasites than C. lahillei. Only 26.9% (21) of the 78 flies infected developed filariae. The highest number of L3 larvae found in an individual fly was one and this was in the head. Development occurred to the L3 stage by PID 7, but the presence of few L3 larvae compared to microfilariae and L2 larvae on PID 8 indicated that the cycle was not synchronous. Of the mean (log +1) microfilarial intake of 49.6 only a mean (log + 1) of 0.04 (0.8%) L3 larvae developed. Despite low biting densities five C. paraensis were experimentally infected and survived to PID 7 - two flies each contained an L3 larva of M. ozzardi in the head. Similarly, only one S. dinellii of eight experimentally infected contained filariae, five L2 stage larvae. Natural infections of flies with filariae were as follows. Of 263 C. lahillei biting man two specimens each contained an L1 larva giving a natural infection rate with filariae of 0.7% and 1.03% (one fly with a single L1 larva) of 97 S. exiguum was infected. DISCUSSION During the selection of a volunteer for the experimental infection study four of the 11 individuals treated two years ago with ivermectin in December 1998 (Table II) were again positive for microfilariae of M. ozzardi in the peripheral blood. Little is known about the effects of ivermectin on this parasite (Campbell 1991), except that Nutman et al. (1987) in a comprehensive study on one infected individual observed that microfilariae had disappeared from the blood for a nine month period following a single treatment with ivermectin (140 µg/kg body weight). It is probable that the drug had suppressed production of microfilariae for this period but not killed the female worm, as occurs in onchocerciasis. Ivermectin is an efficient microfilaricide for O. volvulus but has to be administered at least once annually (150 µg/kg body weight) for many years because new microfilariae produced by female worms will appear if only a single dose is given (WHO 1995). It is not known exactly how many ceratopo-gonid and simuliid species occur in the area of Finca Normenta because no comprehensive seasonal survey has been made. Previous works in this region of Argentina (S Coscarón, unpub. data, Romaña & Wygodzinsky 1950) indicate that S. dinellii bites man in greater numbers in the wet season and that the anthropophilic species, S. jujuyense Paterson & Shannon and the zoophilic species S. lahillei (Paterson and Shannon) are also present. The data presented clearly show that M. ozzardi microfilariae will develop to maturity in C. lahillei, C. paraensis and S. exiguum. The presence of L2 larvae in S. dinellii on PID 5 may have resulted from the experimental infection with M. ozzardi, but could equally have been the result of the fly having had a previous blood meal on another animal that was infected with another filarial species; both O. cervicalis (Roux et al. 1984) and O. gutturosa (Mancebo & Roux 1986) have been recorded in horses and cattle respectively in Argentina. S. dinellii was discounted as a vector in Tucumán province to the north of Finca Normenta by Romaña and Wygodzinsky (1950) because of the failure of microfilariae to develop beyond this stage in flies that had fed on a volunteer infected with M. ozzardi. From our data C. lahillei is clearly a vector in the area because a high proportion of microfilariae developed to infective larvae in the experimental infection and on PID 3 the infective larvae present must have developed from an infection already carried by the fly when it received its experimental infective feed. C. paraensis is also probably a vector. Despite ingesting 15 times more microfilariae than C. lahillei development of the parasite in S. exiguum was poor, but did continue until the L3 stage. Consequently, S. exiguum probably sporadically transmits the parasite in this locality. Only L1 larvae were recorded from naturally infected flies and so it was not possible to identify them to species. Only sufficient data are available for C. lahillei and S. exiguum for comparisons with other ceratopogonid and simuliid vectors of M. ozzardi in Latin America. Our data have shown that in Argentina C. lahillei is an equally efficient host to M. ozzardi as C. furens in Haiti (Lowrie & Raccurt 1981) and C. phlebotomus in Trinidad (Nathan 1981) in terms of infection rates, rapidity of the development cycle and high production of infective L3 larvae from ingested microfilariae. However, not all Culicoides species are efficient hosts to M. ozzardi. C. insinuatus was a less efficient host to this filaria in Colombia (Tidwell & Tidwell 1982), while Lowrie and Raccurt (1981) and Raccurt and Lowrie (1981) showed C. furens to be a more effective host than C. barbosai and Leptoconops bequaerti to be the least effective in Haiti. S. exiguum in Finca Normenta was a relatively poor host to M. ozzardi because of low percentage development of microfilariae to infective larvae and an asynchronous development cycle. This was not the case in Amazonian simuliid species (S. amazonicum, S. argentiscutum and S. oyapockense s.l. [as Simulium sp., S. minusculum, S. sanchezi], reviewed in Yarzábal et al. (1985), which were all more effective hosts in terms of these parameters. It has been shown by Shelley et al. (1987) that S. oyapockense s.l. in the Brazilian Amazon is a relatively poor host to O. volvulus because of the damage caused by its cibarial teeth to microfilariae during ingestion, and that this was a major factor in determining vectorial competence in vector simuliid species in Latin America (Shelley 1988, 1994). Generally, species with unarmed cibaria are more effective hosts to O. volvulus than those with armed cibaria and the same must be true for M. ozzardi. All the species cited in Yarzábal et al. (1985) possess cibarial teeth, whereas S. exiguum at Finca Normenta does not. This would therefore suggest that this simuliid population is a poor host to M. ozzardi because of rapid formation of the peritrophic membrane or because of an inhibitor in the thoracic musculature as is seen in some simuliid species that transmit O. volvulus in Africa and Latin America (Shelley 1988, 1994). Host capacity variations also occur at the level of cytospecies. S. exiguum is a species complex in which individual cytospecies show different capacities to act as hosts to O. volvulus in Ecuador (Shelley et al. 1990) and in which some populations (Venezuela) are actually poor hosts (Shelley 1994). Various data are now available that further refute the theory that two strains of M. ozzardi occur, the Caribbean with a ceratopogonid vector and the South American with a simuliid vector. Tidwell and Tidwell (1982) showed that S. amazonicum, S. argentiscutum and C. insinuatus occurred sympatrically in the Colombian Amazon and that all three acted as intermediate hosts to M. ozzardi, but the ceratopogonid species was less competent as a host than the two simuliid species. Similar findings on these three species were reported by Shelley et al. (1980) from the Brazilian Amazon, except that none of the few C. insinuatus experimentally infected with microfilariae of M. ozzardi produced developing parasites. The evidence that we present here from the Argentina focus shows that species from both families are able to host this parasite but that ceratopogonids are clearly the more effective hosts. Studies on the parasite itself show that populations from Haiti and the Colombian Amazon are almost morphologically identical (Kozek & Raccurt 1983) and that the DNA of the Argentine population from Finca Normenta is identical with that from the Brazilian Amazon (R Morales Hojas, pers. comm.). On the existing evidence, it is therefore more probable that M. ozzardi is a widespread monomorphic species in Latin America that may be transmitted by ceratopogonid and/or simuliid species to varying degrees of competence depending on the vector population concerned. ACKNOWLEDGEMENTS To Dr G Safarov and Dr J Arcaide of the Fraile Pintado Hospital for valuable orientation and use of laboratory facilities respectively; Dr G Spinelli for identification of the ceratopogonids; to Mr H Suligoy for assistance during field work and to Miss A Sanders for assistance in dissecting some of the material. Mr M Fritz kindly provided transport in the field. REFERENCES
[ Medline ] [ Medline ] [ Medline ] [ Medline ] Copyright 2001 Fundacao Oswaldo Cruz Fiocruz The following images related to this document are available:Photo images[oc01069t4.jpg] [oc01069t3.jpg] [oc01069t2.jpg] [oc01069t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}