 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(4) Mar. 2001, pp. 549-553 Isotype Specific Immune Responses in Murine Experimental Toxocariasis C Cuéllar/+, S Fenoy*, C del Águila*, JL Guillén Departamento de Parasitología,
Facultad de Farmacia, Universidad Complutense, 28040-Madrid, España *Facultad
de CC Experimentales y Técnicas, Universidad San Pablo CEU, Madrid, España Received 11 July 2000
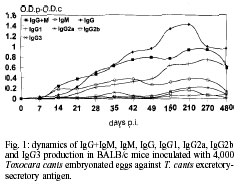
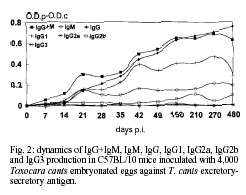
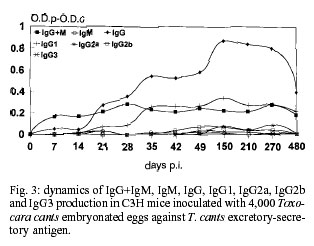
Code Number: oc01084 In this work, a murine experimental model of toxocariasis has been developed in BALB/c, C57BL/10 and C3H murine strains orally inoculated with 4,000 Toxocara canis embryonated eggs, in order to investigate the isotype-specific immune responses against excretory-secretory antigens from larvae. T. canis specific IgG+M, IgM, IgG, IgA, IgG1, IgG2a and IgG3 were tested by ELISA. The dynamics of the specific immunoglobulins (IgG+IgM) production showed a contrasting profile regarding the murine strain. Conversely to the results obtained with the IgM isotype, the IgG antibody class showed similar patterns to those obtained with IgG+IgM antibodies, only in the case of the BALB/c strain, being different and much higher than the obtained with IgG+IgM antibodies, when the C3H murine strain was used. The antibodies IgG+IgM tested in BALB/c and C57BL/10 were both of the IgM and IgG isotypes. Conversely, in the C3H strain only IgG specific antibody levels were detected. The IgG1 subclass responses showed a similar profile in the three murine strains studied, with high values in BALB/c, as in the case of the IgG responses. Key words: Toxocara canis - BALB/c - C57BL/10 - C3H - ELISA - immunoglobulins Visceral larva migrans (VLM) is a clinical syndrome caused by the migration of nematode larvae through the extraintestinal tissues of man (Beaver 1969). Although several nematodes have been reported to produce VLM, Toxocara canis appears to be the primary causative agent (Glickman et al. 1979). The signs and symptoms of human toxo-cariasis have been described by Barriga (1988), who grouped them in diminishing order according to diagnostic significance. Peripheral eosinophilia, leukocytosis, hypergammaglobulinemia, hyperiso-hemagglutinemia, anaemia and hypoalbuminemia were the principal laboratory findings in VLM cases, and hepatomegaly, respiratory signs, fever, malnutrition, digestive signs, asthenia, nervous signs, spleenomegaly and anorexia were the more frequent symptoms. The patient with VLM is usually a child, one to five years old, with a history of pica and contact with pets (Cypess et al. 1977, Overgaauw 1997). The murine model has been used as the most adequate for the study of human infection by T. canis. In this paratenic host there is survival but no development (Beaver 1969). The migratory route in the mouse includes two phases: a visceral phase, during the first week post-infection (p.i.), when larvae mainly reach the liver and lungs (hepato-pulmonary phase); afterwards, larvae migrate throughout the body and accumulate principally in the carcass and in the brain (myotropic-neurotropic phase) (Abo-Shehada & Herbert 1984). In a previous work (Guillén et al. 1990), comparison was made of the migratory behaviour of T. canis larvae in two inbred strains of mice (BALB/c and C57BL/10), analyzing liver, lungs, viscera (heart, kidney and spleen), muscle of hindfoot, carcass and brain. In BALB/c mice, migratory capacity was faster than in C57BL/10 and with no return from the brain, while in the latter strain, larvae went back to finally accumulate in the liver (day 63 p.i.). Afterwards the ELISA method, using excretory-secretory (ES) products, was used to determine the development of circulating antibodies in BALB/c and C57BL/10 mice. When mice inoculated with T. canis embryonated eggs were tested the peak was detectable at the 9th week in BALB/c and at 7th in C57BL/10 (Cuéllar et al. 1990). In order to investigate the isotype-specific immune responses, a murine experimental model of toxocariasis has been developed in BALB/c, C57BL/10 and C3H mice orally inoculated with 4,000 T. canis embryonated eggs. The aim of this study was to improve our knowledge of the host-parasite relationship in order to elucidate the immunopathological mechanisms of toxocariasis. MATERIALS AND METHODS T. canis eggs - T. canis eggs were obtained by dissection of gravid females, resulting from a natural infection. After purification, they were embryonated under sterile conditions at 37ºC and preserved at 4ºC until use (Guillén et al. 1986). Production of ES antigen from T. canis larvae - T. canis larvae were obtained from embryonated eggs submitted to the action of a mixture of equal parts of sodium hypochloride and sodium hydroxide at 2% to remove the external shell, ending the hatching by mechanical treatment with sterile glass beads. To obtain the antigen, a modification of the de Savigny (1975) method was used by which the larvae were cultivated in Eagle's Essential Minimum Medium with Earle's Salts. Each tube contained 5 ml of medium with a larval concentration of 1X104/ml. Supernatants were collected weekly. After dialyzing against PBS and concentrating in Amicon YM-10, its protein content was estimated by the Bradford (1976) method and finally frozen at -20oC until use (Águila et al. 1987, Cuéllar et al. 1995). Experimental animals - Ten mice from the BALB/c, C57BL/10 and C3H strains, respectively were used. Adequate measures were taken to minimise pain or discomfort in experimental animals. Experiments were carried out in accordance with the European Communities Council Directive of 24 November 1986 (86/609/EEC). Inoculation protocol - Groups of mice were inoculated by oral administration with gastric tubing of dose of 1,000 embryonated T. canis eggs per animal weekly, during four weeks. Serum - Animals were bled under ether anaesthesia, by the retroorbital plexus on days 0, 7, 14, 21, 28, 35, 42, 49, 150, 210, 270 and 480 p.i. After bleeding, blood samples from each group of animals were pooled and centrifuged to obtain sera. Specific antibody levels - Specific antibodies levels were measured by ELISA. The 96 well microtitre plates (Nunc-Immuno Plate Maxi SorpTM ) were coated overnight at 4ºC by the addition of 1 µg/ml of larval ES antigen diluted in a carbonated buffer to 0.1M at pH 9.6. Several wells were kept uncoated as control for nonspecific reactions (Cuéllar et al. 1992). After washing three times with 0.05% PBS-Tween 20 (PBS-Tween), blocking was carried out by adding 250 µl per well of 0.1% BSA in PBS, incubating for 1 h at 37ºC. After washing, 100 µl of serum samples diluted 1/100 in PBS-Tween, 0.1% BSA were added in duplicate and incubated at 37oC for 2 h. As negative control, sera from non-infected mice were used (all animals were bled at day 0). Once the plates were washed, 100 µl per well of goat affinity isolated, horseradish peroxidase conjugated specific to mouse IgG + IgM (H+L), IgM (µ), IgG (g), IgG1(g1), IgG2a (g2a), IgG2b (g2b), IgG3 (g3) and IgA (a) (CALTAG Laboratories, San Francisco, CA), at the appropriate dilution in PBS-Tween, 0.1% BSA, were added and incubated for 1 h at 37ºC. After washing, 100 µl per well of substrate (o-phenylene-diamine; SIGMA) were added at 0.04% in a phosphate-citrate buffer (pH 5.0) with 0.04% hydrogen peroxide. The reaction was stopped with 3N sulphuric acid and the plates were read at 492 nm. Results were expressed as O.D.p-O.D.c indexes by subtracting the mean O.D. of the control from the mean O.D. of the test sera once the non-specific reaction with the BSA used in the blocking was subtracted. RESULTS Dynamics of antibody responses in BALB/c mice - Antibodies (IgG+IgM) were first detected between 14 and 21 days p.i., peaking at day 210 p.i., remaining at a high level until the end of the experiment. Specific antibodies of the IgM isotype showed a rapid production with a great response between days 42 and 210 p.i., followed by a decline by the end of the experiment. Levels of antibodies were high when the IgG isotype was tested, showing maximum values between days 105 and 210 p.i., and remaining at very high levels for the remainder of the experiment. The production of the IgG1 subclass was first detected after day 14 p.i. with high levels for the rest of the experiment. Very low responses were observed when the IgG2a and IgG2b subclasses were investigated, with indexes never greater than 0.2. In the same manner, very low levels of IgG3 were present (Fig. 1). Finally, IgA also showed very low levels. Dynamics of antibody responses in C57BL/10 mice - Specific antibodies (IgG+IgM) with indexes greater than 0.2 were detected after 21 days p.i., peaking at day 210 p.i. and remaining between 0.4 and 0.6 at the end of the experiment. The IgM isotype showed oscillations remaining between 0.1 and 0.2 during the entire experiment. Specific antibodies of the IgG isotype showed detectable levels after 21 days p.i., peaking at the end of the experiment. When the IgG1 subclass was tested, antibody levels increased after 21 days p.i. Very low responses (lower than 0.1) were observed when IgG2a, IgG2b and IgG3 subclasses were investigated (Fig. 2). Indexes never greater than 0.015 were observed when IgA responses were tested. Dynamics of antibody responses in C3H mice - Specific antibodies (IgG+IgM) with indexes greater than 0.2 were only detected by day 28 p.i., remaining at low levels during the whole experiment. Indexes never greater than 0.1 were observed when the IgM isotype was tested. Antibodies of the IgG isotype were first detected after 21 days p.i., with a peak at day 150 p.i. Similar profiles were observed when IgG1 responses were tested. Indexes never greater than 0.1 were observed when IgG2a, IgG2b, IgG3 and IgA responses were studied (Fig. 3). DISCUSSION The dynamics of the specific immunoglobulins (IgG+IgM) production in BALB/c, C57BL/10 and C3H murine strains orally inoculated with 4,000 T. canis embryonated eggs showed a contrasting profile regarding the murine strain. The results suggest that the C3H strain is a low-responder to ES products released by the larvae during the infection, otherwise the larvae could not be capable of surviving in this non-susceptible strain. On the other hand, in this strain, the inoculum of 4,000 embryonated eggs could have been a high infective dose inducing a partial immunosuppression of mice, as observed by Boroskova et al. (1998) and Dvoroznakova et al. (1999) by inhition of the proliferative response of spleenic T and B cells to polyclonal activators. The IgG antibody class showed similar patterns to those obtained with IgG+IgM antibodies, only in the case of the BALB/c strain, being different when the C3H murine strain was used. The antibodies IgG+IgM tested in BALB/c and C57BL/10 were both of the IgM and IgG isotypes. Thymus-independent antigens are present in the ES products released by the living larvae (Maizels et al. 1987) and their products could be generated only in high and medium susceptible strains. The IgG responses observed in the C3H strain could, probably due to products released from the dead larva and their products, be recognized by ES antigen. The IgG1 subclass responses showed a similar profile in the three murine strains studied, with high values in BALB/c, as in the case of the IgG responses. This results might indicate that somatic products released from the dead larvae are responsible for IgG1 production observed in the C3H strain. However, Epe et al. (1994) examined the behaviour and pathogenicity of second-stage larva (L2) of T. canis in different mouse strains. The highest larval counts in the brain of all strains were found in BALB mice. The percentage of eosinophils in the blood of BALB mice increased after the 8th week p.i., whereas it decreased in the other strains, with histological and pathophysiological changes of a lesser extent in this strain than in the others. In mice of the strains C3H and C57BL, deaths occurred from the 4th week p.i. onward, and the number of larvae found in their brains were lower than those observed in BALB. Koizumi and Hayakawa (1982) showed that the number of larvae recovered from the liver of BALB/c, C57BL/6 and C3H/He mice infected by T. canis was significantly higher in BALB/c mice. Secretion of IgG2a is enhanced by Th1-dependent IFN-g (Snapper et al. 1988). The presence of both IgG1 and IgG2a isotypes in the BALB/c strain could suggest that both Th1 and Th2 cells are active, although with a Th2-dominant response during the T. canis infection. This BALB/c strain seems to be a high-responder murine strain, conversely to the results observed in the apparently medium- and low-responder C57BL/10 and C3H strains, where only Th2 cells could be active. These results are in accordance with the features cited by Else and Finkelman (1998) who observed that Th1 type responses are associated with susceptibility to Trichuris muris infection. In BALB/c mice, migratory capacity of T. canis larvae is faster than in C57BL/10 and with no return from the brain, while in the later strain, larvae go back to finally accumulate in the liver (Guillén et al. 1990). Results from Guillén et al. (1990), Bardón et al. (1994), Sugane et al. (1996) and Dent et al. (1999), confirmed the high susceptibility of the BALB/c strain in which both Th1 and Th2 cells are active with production of specific IgG1 and IgG2a isotypes. We have previously reported that sera from BALB/c mice orally inoculated with 1,000 T. canis embryonated eggs showed detectable specific immune complexes after the first week p.i., increasing progresively until the 6th week p.i. and remaining at a high level during the entire experiment (eight weeks), with a similar profile as that observed when antibodies to the ES antigen were tested. This fact suggests that the larvae were alive during this time in this susceptible strain with production of ES substances which are responsible for the formation of immune complexes. In the case of larval destruction or immobilization the rapid clearance of specific immune complexes results in their almost complete disappearance from the circulation and the small amounts of ES substances released by the cuticle of the destroyed larvae is insufficient to alter this balance (Bardón et al. 1995). From the results presented, it can be concluded that humoral immune responses against T. canis antigens are different depending on individual characteristics. REFERENCES
[ Medline ] [ Medline ] Copyright 2001 Fundacao Oswaldo Cruz Fiocruz The following images related to this document are available:Photo images[oc01084f2.jpg] [oc01084f3.jpg] [oc01084f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}